

Самец певчего воробья в Prospect Park, Нью-Йорк, Бруклин
Борьба за существование гибельна для обоих сторон
Содержание
Читая Дарвина, обратил внимание на малоизвестную идею Томаса Роберта Мальтуса, им воспринятую и легшую в структуру теории эволюции путём естественного отбора (в отличие от известной пошлости что «пряников сладких всегда не хватает на всех», обычно к тому же неверной: размножается больше, чем может выжить не из-за нехватки корма, а из-за того, что при избыточном размножении ты сам оказываешься слишком лёгким кормом даже для неспецифических хищников/паразитов. По крайней мере, до экспансии человеческого хозяйства, отчуждающего себе всё большую часть продукции биосферы, было именно так.).

Это представление, что отбор связан с борьбой за существование так, что непосредственно во время неё организмы чувствуют себя хуже, чем без борьбы. Кроме снижения численности проигравших и перераспределения ресурсов между ними и выигравшими в борьбе за существование, в отбор вносит вклад и сам факт борьбы: раны и физиологические проблемы получают и выигравшие, и проигравшие, конкурентный голод не тётка для обоих. Поэтому во время борьбы обе «стороны» размножаются хуже, чем без или после неё.
В том числе вследствие обоюдной «психологической дуэли», связанной с фактом обнаружения присутствия хищника по его крикам, следам, запаховым или мочевым меткам и пр. Для хищника не менее травматичен факт обнаружения его жертвой, он стрессируется, и часто после этого отказывается от охоты: о том и другом дальше рассказывается в посте.
Вот недавний пример: перед и во время драк молодых фазанов Phasianus colchicus с установившимся «порядком клевания» (т.е. лестницей иерархии) стресс испытывают обе особи, независимо от статуса и успеха в прошлых стычках, что проявляется в снижении температуры головы (кровь уходит в «ядро» корпуса, обычное следствие стресса). А после драки стресс проходит у обоих, голова теплеет, независимо от того, что выиграл, кто проиграл. Самки стрессируются сильнее, судя по «холодению» головы (Knoch et al., 2022).
Отсюда следует, что ни один признак дарвиновых вьюрков, или домашних животных, или иных модельных объектов эволюционной теории не выведен из-под отбора, все они так или иначе «действуют» в борьбе за существование, но вклад только некоторых выделен и решающ.
Хищник-жертва: психологическая «дуэль»
Я писал, как запаховые метки хищника подавляюще воздействуют на потенциальных жертв, вплоть до сдвига соотношения полов в их потомстве и гормональных сдвигов у них самих. Сейчас выясняется, что также действуют и «акустические метки»: крики и т.п. звуки жизнедеятельности. Восприятие жертвами криков хищников влияет на их репродукцию, они меньше «вкладываются» в производство потомства, с соответствующими последствиями для популяции. Отсюда потенциальные жертвы «оценивают» риск не только на основании непосредственно испытанных нападений, заканчивающихся ранением и гибелью, но в первую очередь по сигналам о постоянном присутствии опасности в районе активности размножающейся пары. Но поскольку первое очень трудно отделить от второго, «психологический» эффект присутствия хищника и его влияние на выживаемость молодых / на воспроизводство популяции до сих пор не исследованы «в чистом виде».
Этот пробел восполняет исследование Liana Y. Zanette et al. (2011), выполненное на одной из американских овсянок – певчем воробье Melospiza melodia. Оказалось, что в ответ на простое озвучивание криками хищников (причём гнёзд птиц защищaлись «инженерными методами» и на деле они были в безопасности) самки овсянок, участвующие в размножении, сокращали репродуктивный выход примерно на 40%.
Полевой опыт ставился на диких, свободноживущих самках M.melodia, строящих гнёзда и выращивающих потомство; в ходе его направленно устраняли возможность реального нападения хищника и проигрывали в течение всего сезона размножения: опытной группе – крики хищника, контрольной – крики безобидных видов. Певчий воробей – широкораспространённый и превосходно изученный вид, гнездящийся в кустарнике, с открытыми чашевидными гнёздами. В районе исследований (несколько малых островов из группы Gulf Islands, Британская Колумбия, Канада) он обитает круглогодично и даёт 2 кладки в год, теряя по крайней мере половину возможного потомства от разорения гнёзд хищниками.

Гнездо певчего воробья
Авторы прямое хищничество исключали, защищая каждое из гнёзд оградой под током «снизу» и сеткой «сверху». Исключение подтверждалось посредством непрерывного видеослежения за гнёздами в течение всего сезона размножения, позволявшего быть уверенным в судьбе каждого яйца и птенца.
Привыкнув к экспериментальным манипуляциям, птицы за гранью дружеских штыков под «колпаком безопасности» размножались не хуже, чем раньше. Это позволило перейти ко второй части опыта – устранив реальную опасность, оценить чисто психологический эффект восприятия риска хищничества по сигналам последнего. Для этого собрали две серии из криков / звуков разных видов, в первой – хищники (енот, ястреб, ворона, воловьи птицы – гнездовые паразиты), во второй неопасные виды (тюлени, гуси, золотые дятлы, гагары, колибри). Каждому из криков хищника был сопоставлен акустически сходный с ним крик безобидного вида (например, карканье ворона и гоготание гуся). Действительно, последующий анализ показал, что между обоими сериями стимулирующего материала нет различий в частотных характеристиках; иными словами, если чем первая серия и отличается от второй, так это наличием или отсутствием информации о потенциальной опасности, а не силой/интенсивностью акустической стимуляции как таковой.
Обе серии звуков не отличались ни по расстоянию от гнезда, на которой находился воспроизводящий их динамик, ни по естественной частоте встречаемости разных криков (или звуков) хищников в природе. Те и другие крики проигрывали каждые несколько минут, все 24 ч в день, в режиме «4 дня птиц озвучивают – 4 дня нет» в течение всего 130-дневного сезона размножения, на общей площади 130 га.
Будущие гнездовые территории учитывали и картировали начиная с марта 2010 г.; затем фиксировали все случаи гнездования самок-владелиц в течение сезона размножения. Каждую из 2-х серий криков проигрывали 12 самкам; конкретные «точки проигрывания» выбирали так, чтобы охватить возможно большую территорию внутри острова, и включить место наблюдения и индивидуальность самок как случайные факторы во всех произведённых анализах.
Динамики размещали через каждые 0,4 га, проигрывание криков начали 15 марта, за несколько недель до появления первого яйца в кладках. Помимо репродуктивного выхода – общего числа слётков, произведённых за год, опытных и контрольных птиц сравнивали по следующим характеристикам репродукции:
- масса яиц и птенцов (влияющая на выживание, как в гнезде, так и в первый год жизни);
— чувствительность птенцов к холодовому стрессу (=сформированность у них терморегуляторных реакций);
— мерили температуру тела через 10 мин после слёта самки с гнезда;
— 4 поведенческих характеристики, отражающих влияние на использование местообитаний (сдвиг типичного места размещения гнёзд), уровень бдительности (измерен по дистанции взлёта – расстояние, на котором самка взлетает с гнезда при приближении наблюдателя), посещаемость гнезда (по длительности интервалов насиживания, между которыми самка оставляет гнездо) и кормление (число приносов корма в час за период выкармливания).
Что получилось? Самки, озвучиваемые криками хищников, произвели за сезон на 40% меньше потомства, чем контрольные (рис.1А), ибо они
- клали меньше яиц (рис.1B)
— у них большая доля яиц повреждалась и гибла при насиживании (что обычно бывает, если родитель слишком часто & неаккуратно переворачивает насиживаемые яйца, а это связано с аномально повышенным уровнем беспокойства), рис.1С;
— в период выкармливания у них погибал больший % птенцов, в основном от недокорма (рис.1D).
См. рисунок 1
Рисунок 2 показывает тот же процесс в динамике, от апреля к июню; видно, что различия в репродуктивном выходе между опытной и контрольной группой растут с каждой стадией репродуктивного цикла (130 дней).
Приведены лишь успешные гнёзда, все самки в обоих группах дали 2 кладки, а некоторые – 2 из озвучиваемых криками хищников, 3 – из озвучиваемых мирными видами – дали три кладки. Звёздочки — значимые различия.
Обратите внимание, что эффект уменьшения репродуктивного выхода от гибели яиц/птенцов от «менее тщательной» заботы о потомстве в опытной группе вполне сравним с эффектом уменьшения кладки и даже перекрывает его, т.е. «психологическое» воздействие хищника столь же сильное, как прямое.
Хотя различия в числе яиц были более выражены в первых кладках, по доле погибших птенцов – во вторых, но в целом совместное влияние факторов «серия криков * № кладки» оказалось незначимым.
Далее, самки, на которых воздействуют криком хищника, размножаются по принципу «лучше меньше, да лучше»: хоть у них больше яиц, но средняя масса яйца повышена (рис.3А), их выводок весит меньше (рис.3В), а птенцы более чувствительны к терморегуляторному стрессу (рис.3С).
Рисунок 3
Опытная и контрольная группа также сильно отличаются по поведенческим характеристикам.
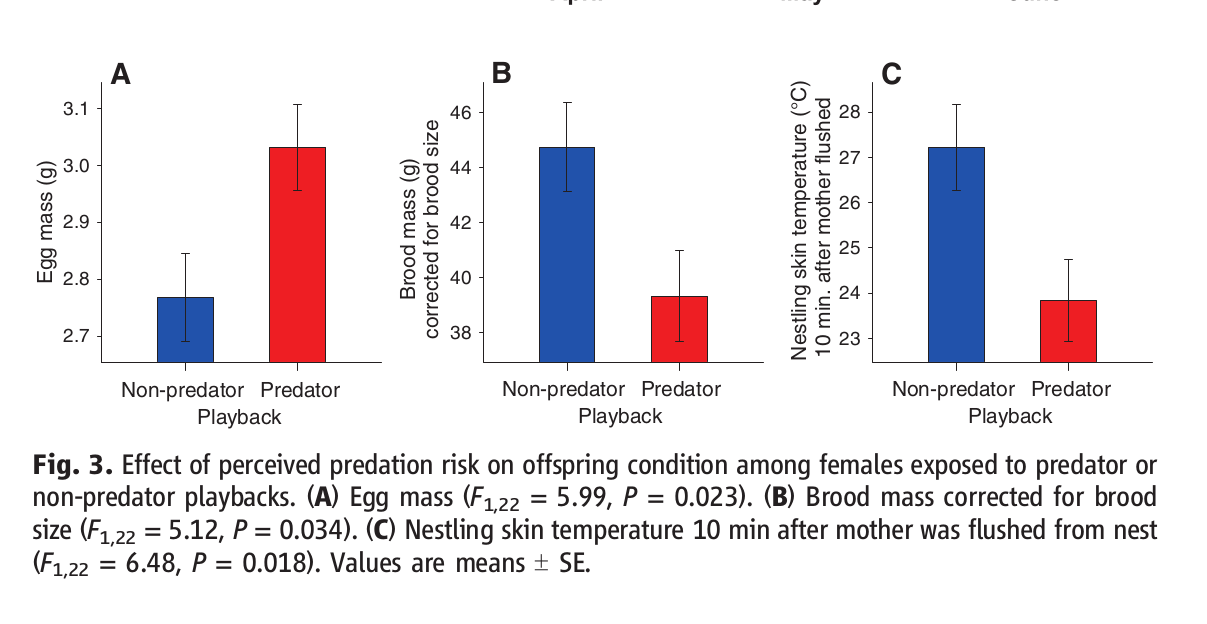 Во-первых, есть явная разница в выборе гнездовых местообитаний. Самки, озвучиваемые криками хищников, строят гнёзда в самых плотных и максимально колючих из кустарниковых зарослей, имеющихся на данной территории (рис.4А), они много более осторожны (что видно по возрастанию дистанции вспугивания, рис.4В), интенсивность кормления птенцов у них значимо ниже (рис.4D), они проводят меньше времени на гнезде и больше времени вне гнезда, чем самки контрольной группы (рис.4С).
Во-первых, есть явная разница в выборе гнездовых местообитаний. Самки, озвучиваемые криками хищников, строят гнёзда в самых плотных и максимально колючих из кустарниковых зарослей, имеющихся на данной территории (рис.4А), они много более осторожны (что видно по возрастанию дистанции вспугивания, рис.4В), интенсивность кормления птенцов у них значимо ниже (рис.4D), они проводят меньше времени на гнезде и больше времени вне гнезда, чем самки контрольной группы (рис.4С).
См. рисунок 4
 Реально «психологический» эффект воздействия хищника на темпы роста популяции ещё больше, чем зафиксированное уменьшение репродуктивного выхода на 40%, ибо включает опосредованные и отсроченные эффекты воздействия «детства в опасности» на молодых птиц в первую осень-зиму их жизни и далее, в сезон размножения. Недокорм (в степени, зафиксированной у птенцов в опытной группе), вредно влияет на физиологию молодых и особенно – на развитие мозга, что отрицательно сказывается на выживаемости. Да и стресс, который явно испытывают самки, направленно деформирует поведение молодых, и не на пользу.
Реально «психологический» эффект воздействия хищника на темпы роста популяции ещё больше, чем зафиксированное уменьшение репродуктивного выхода на 40%, ибо включает опосредованные и отсроченные эффекты воздействия «детства в опасности» на молодых птиц в первую осень-зиму их жизни и далее, в сезон размножения. Недокорм (в степени, зафиксированной у птенцов в опытной группе), вредно влияет на физиологию молодых и особенно – на развитие мозга, что отрицательно сказывается на выживаемости. Да и стресс, который явно испытывают самки, направленно деформирует поведение молодых, и не на пользу.
Таким образом, было показано, что «подавляющий» эффект воздействия популяции хищников на рост численности популяции жертв сильно больше эффекта прямого изъятия. Ведь все 3 демографических измерения «психологического» воздействия хищника (рис.1В-D) реально влияют на репродуктивный выход популяции в те же годы, когда реальное воздействие хищника вполне присутствует. Более того, величина эффектов рис.1В-D сравнима с аналогичными изменениями в ответ на реальное возрастание риска хищничества, а результаты опытов с многолетней подкормкой певчих воробьёв показывают, что эффекты реакций на хищника вполне устойчивы к вариациям в обеспеченности кормом. Т.е. они представляют собой один из аспектов жизненной стратегии вида.
Результаты проделанных экспериментов показывают, что эффекты субъективного восприятия видом-жертвой уровня риска сами по себе – достаточно мощное средство для оценки скорости роста популяции жертв в дикой природе. Аналогичные результаты были получены на млекопитающих: в исследованиях системы «лось – волк», снежных коз, дюгоней и акул, горлиц Zenaida asiatica их хищников — краснохвостых канюков Buteo jamaicensis (см. также табл.2 данной статьи с перечнем видов, изученных в плане реакции на сигналы присутствия хищников). К слову, это говорит о символизации внешнего мира позвоночных, об их, так сказать, примитивненьком логосе, ограниченном рамками видового Umwelt’a – сигналы о хищнике, знаки его присутствия в местообитании, управляют динамикой популяций не хуже, а то и лучше, чем реальные нападения оного.
Эти и некоторые другие данные показывают, что оценка риска хищничества в популяции жертв происходит не только по первичным, но и по вторичным сигналам присутствия хищников в их среде обитания. Первые – это собственно частота гибели/поранения особи при нападениях, второе – крики, запаховые, следовые метки и т.п. «следы присутствия» опасности вокруг индивида. Их пространственная и временная плотность сигнализирует о необходимости «сдвинуть реле» между вкладом в воспроизводство (с развертыванием соответствующего поведения в виде ухаживания, охраны территории, образования / консолидации пары, с последующей заботой о потомстве).
Я уже писал, что в жизни большинства позвоночных присутствует фундаментальный конфликт интересов (по-английски trade-off) между вкладом в воспроизводство следующего поколения и вкладом в удовлетворение собственных витальных потребностей, т.е. в себя любимого. Причём этот компромисс в каждом следующем сезоне размножения надо достигать заново, ориентируясь на сравнение сигналов из внешней среды, экологической и социальной, с собственным состоянием и «силами» животного, куда входят физические силы, уровень ресурсообеспеченности, уровень агрессивности, уровень брачного возбуждения, и уровень стресса.
{Рост последнего сигнализирует о выходе интенсивности средовых воздействий за оптимальный уровень, о том что требуются напряжение и мобилизация сил, на которую готовы не все, а только лишь «сильные» и «успешные» особи. Сам вид воздействия тут не очень существенен – и увеличение риска хищничества, и рост напряжённости социальных контактов, и экстремальные условия внешней среды, погодные или кормовые, равно опосредуются увеличением концентрации гормонов стресса. Как это влияет на жизненные циклы особей, какие вызывает отклики в виде физиологических/поведенческих изменений, хорошо рассказано в обзорной статье Е.А.Новикова и М.П.Мошкина (2009) «Роль стресса в модификации поведенческих программ».}.
Первое предполагает активное участие в репродукции, высокие затраты как на само производство яиц и детёнышей, так и на те формы брачной активности (и шире, всей социальной активности, вроде занятия территорий, их охраны и рекламирования, поддержания определённого рода иерархии в группах и пр.), без которых размножиться не удаётся. Как я уже писал, самую «эгоистическую» потребность в максимизации итоговой приспособленности не удовлетворить в одиночку, надо взаимодействовать с другими индивидами, при этом «укладывая» собственную активность в видоспецифические формы демонстраций, а поддерживаемые структуры отношений — в видовую норму социальной организации. Выход первого и второго за пределы видовой нормы делает все вложенные усилия напрасными.
Первое, с одной стороны, предполагает значительный выигрыш. С другой, оно связано с существенно большим риском, ибо всякое активное участие в размножении и особенно в той социальной активности, без которой последнее невозможно и неэффективно, увеличивает риск (в том числе подверженность гибели от хищников) и снижает иммунитет, увеличивая подверженность болезням и стрессам. Подробней об этом см. здесь и здесь.
Второе же – выбор в пользу больших усилий в самосохранении, нежели в репродукции, означает отказ от выигрыша или существенно меньший выигрыш, ибо животное «пропускает ход», не участвуя в данном сезоне размножения или участвуя с существенно меньшей успешностью, чем «средние» особи в популяции. Зато эти особи избегают высоких затрат и высокого риска, и к следующему сезону размножения имеют шансы прийти в существенно лучшем состоянии, нежели индивиды, сделавшие выбор противоположного рода.
«На одном полюсе градиента «качества» (без определения, кто выше, кто ниже), особи, выигрывающие в социальной конкуренции: в каждый данный момент они готовы максимально развернуть агрессию или ухаживание, даже при паразитарных стрессах, когда другие особи вкладываются в иммунный ответ. Выигрывая в социальной конкуренции, они проигрывают в подверженности средовым стрессам – особенно в экологически неблагоприятные годы. На другом полюсе градиента «качества»: особи, устойчивые к энвиронментальному стрессу, но проигрывающие в социальной конкуренции: неблагоприятные изменения среды у них вызывают хороший иммунный ответ, они легко отказываются от социальной активности ради прекращения преследования доминанта, физиологического ответа на неблагоприятные изменения среды и т.п. См. 1, 2 и 3.» [Источник].
Повторюсь: выбор в пользу одной из возможностей (максимизация выигрыша или минимизация риска) каждый сезон размножения нужно совершать заново, или же достигать оптимального компромисса между ними, структурируя собственное поведение так, чтобы достигались обе возможности, но частично. Соответственно, акты совершения выбора всё сильнее дифференцируют особей в популяции. Условно говоря, особи «сильные», с высокой эффективностью коммуникации и/или с высоким уровнем ресурсообеспеченности выбирают активность, связанную с размножением в ущерб активности, связанной с самосохранением, особи «слабые», сильно стрессированные и/или плохо обеспеченные ресурсами – наоборот. Отсюда возникает существенная поливариантность онтогенезов особей в популяциях позвоночных.
Обе возможности строго альтернативны: животное или идёт на резкое увеличение риска, чтобы развернуть соответствующую последовательность демонстраций до конца и, в случае успеха взаимодействия с соответствующими компаньонами, «взять» долговременный выигрыш, или отказывается от любого увеличения риска, наоборот, всячески минимизирует риск и занимается только активностью самообеспечения. Оно интенсивно кушает, бережёт себя и набирается сил, что позволяет выжить в неблагоприятных условиях, но «пропускает ход» в плане социальной активности и воспроизводства, в надежде на то, что в следующий сезон условия будут лучше и возможностей поучаствовать в размножении будет больше.
Соответственно, поливариантность жизненных стратегий особей в популяции определяется тем, какой % особей в данной популяции выбирает первый вариант (рисковать, но вкладываться в социальные взаимодействия), а какой второй (минимизировать риск, «пропускать ход»). Среди особей первой категории та же поливариантность стратегий определяется тем, до какой степени разные особи готовы идти на риск и продолжать демонстрировать «по полной программе» в ответ на ухудшение условий среды, появление опасности гибели и т.д.? При каком уровне риска и каком ухудшении ресурсообеспеченности либо погодных условий разные особи переходят к ухаживанию или охране территорий «по экономной программе»?
И конечно же, жертва «не безоружна» в психологическом смысле. В исследовании E.L.Shally & D.T.Blumstein (2005), посвящённом эволюции сигналов предупреждения об опасности у разных групп дневных социальных видов грызунов показано, что на первой стадии этой эволюции, когда сигнал представляет собой сигнал состояния, а не ситуации (т.н. сигналы-стимулы), он дезориентирующее действует на хищника. Фактически это в большей степени сигнал хищнику (чем сородичам), что он обнаружен и жертва настороже, волнуется, мобилизуется и пр., что хищника «обескураживает», он стрессируется и отменяет атаку, либо его охотничьи стереотипы «сбоят» от воздействия моббинга жертвы и атака оказывается неудачной.
Как сигнал сородичу (особенно непосредственно не видящему опасность и не боящемуся её) сигнал-стимул достаточно плох, ибо передаёт субъективную степень страха особи, продуцирующей сигнал, а точность реакции на него пропорциональна уровню страха реципиента, т.е. коли сородич не видит и не боится опасности, верно не среагирует на сигнал. Зато хищник обезоружен!
Видимо, потому, что «психологические воздействия» от сигналов присутствия хищника и тревожной вокализации жертвы взаимно уравновешивают друг друга, издавание тревожных криков у сурков и ряда других видов не увеличивает риск погибнуть для кричащей особи, вопреки известному социобиологическому построению. А вот антропогенный или техногенный шум, от людей и работающей техники, дезориентирует особей, затрудняет реакцию на видовые сигналы тревоги и значимо повышает вероятность гибели от тех же хищников сравнительно с природными местообитаниями, где нет этого шума (+тут). Я сам люблю теоретические спекуляции, но лучше уж стены высокого замка теории возводить с постоянно наращиваемых «лесов», образуемых связанными между собой данными эмпирии, нежели заранее выстроенную теорию инкрустировать «парадными примерами» фактов, как шалашник раскладывает украшения перед «беседкой». В первом случае в возведённой конструкции можно жить, в смысле развивать теорию дальше, успешно инкорпорируя в неё новое знание, включая наблюдения и опыты, ранее непредвиденные, во втором получившаяся инсталляция годится только на то, чтобы трахнуть самку, в смысле, создаст некритичных приверженцев данному объяснению. Модность, однако не делает его более состоятельным.
Вернёмся к работе E.L.Shally & D.T.Blumstein (2005). Следующая стадия эволюционного совершенствования сигнальной функции тревожных криков, независимо наступающая в разных филогенетических ветвях грызунов – это превращение тех же самых (или сильно гомологических) демонстраций из сигналов-стимулов в сигналы-символы. Последние указывают на определённые классы опасностей, фактически «именуют» их, они эмансипированы от мотивационной подосновы, т.е. нормально издать специфический крик тревоги и эффективно отреагировать на услышанный крик может и особь, не видящая опасности, не боящаяся её в должной степени и пр., причём реакция будет точно подогнана к данному виду хищника. Иными словами, сигналы – стимулы мощное средство индивидуальной адаптации и хреновое – групповой; обезопасивая данную особь, они плохо передают информацию другим. А вот сигналы-символы – наоборот, и что в самых разных филогенетических ветвях эволюция идёт от первых к вторым, крайне симптоматично.
Источник wolf_kitses
Психологическая «дуэль» обоюдна: сигналы тревоги жертв сдерживают хищников, заставляют отказываться от охоты
«Психологическая дуэль» между хищником и жертвой обоюдна — не только жертва стрессируется сигналами его пребывания, как описано выше; хищник тоже стрессируется, когда его, обнаруженного, окрикивает потенциальная жертва, почему после этого обычно отказывается от охоты. Тем более что успех последней требует незамеченности, особенно у засадников; однако окрикиваемые часто настолько стрессированы, что даже и не пытаются «зайти с другой стороны», снова стать незаметными, сменив место засады и пр., а просто уходят прочь.
Так, обработка тревожными криками мартышек «на леопарда» (у них есть ещё совершенно иначе звучащие «на орла» и «на питона» — действуют именно эти) заставляет леопардов, помеченных ошейниками с радиопередатчиками, разворачиваться и двигаться прочь из данного района. Окрикивание обезьянами пресекает даже спокойный отдых хищника: длительность периодов его отдыха «до» обнаружения распределена равномерно, периодов отдыха «после» сокращена до одного и того же — минимального — интервала. Аналогична реакция на окрикивание немеченных леопардов Panthera pardus: незамеченные группами обезьян продолжают свой отдых ещё достаточно долго, замеченные прерывают его через несколько минут, чем обнаруживают себя. См. данные по 6 видам обезьян леса Таи, Берег Слоновой Кости, двум тонкотелам (Colobus badius, C.polykomos), трём мартышкам (Cercopithecus petaurista, С.diana, C. campbelli) и дымчатому мангобею Cercocebus atys (Zuberbühler et al., 1999).
Этот эффект отсутствует при обработке любыми другими криками данного вида, в том числе и другими тревожными. Другой важный момент: кроме леопарда, на обезьян здесь охотятся шимпанзе Pan troglodytes. В ответ на записи «уханья» шимпанзе обезьяны издают те же тревожные крики, что в ответ на рычание леопарда (видимо, связанные со страхом перед любыми хищными млекопитающими), однако значительно реже. Во втором случае окрикивающие обезьяны активно подходят к источнику звука, в первом же никогда. В отличие от леопарда, шимпанзе этим криком не остановишь, а прямо наоборот, он облегчает им поиск и отлов жертв.
Чем дополнительное подтверждается отсутствие у антропоидов инстинктов, также как у людей, иначе б они «автоматически» реагировали на инстинктивные же сигналы тревоги мартышек и тонкотелов, по образцу леопардов или хищников, окрикиваемых сурками и другими наземными социальными грызунами. Аналогичные данные получены для меченых GPS-ошейниками верветок Chlorocebus pygerythrus и леопардов в Кении. Крики тревоги верветок, услышанные леопардами на дистанции <200 м, заставляли их поворачивать прочь, если обезьяны молчали, кошки продолжали двигаться вперёд. Данные фотоловушек, выставленных на местах ночлега верветок, показывают, что когда обезьяны молчат, леопард может к ним подойти на существенно меньшее расстояние, чем когда они издают тревожные крики. Поэтому попытки охоты происходят в основном ночью, а не утром или вечером, когда интенсивность окрикиваний существенно выше (Isbel, Bidner, 2016). Сходным образом полевые жаворонки Alauda arvensis, заметив опасного хищника — дербника Falco columbarius, начинают петь. Непоющих или малопоющих птиц дербник преследует дольше и ловит чаще, чем поющих интенсивно. Влияния температуры на все перечисленные характеристики не обнаружено. Охотящийся дербник предпочитает крупные скопления жаворонков, почему вероятность быть атакованным в крупной стае растёт, однако успешность охоты не связана с размером стаи (Cresswell, 1994).
Такие же наблюдения сделаны и в Перу над оцелотами Leopardus pardalis, меченными радиофицированными ошейниками. Исследователи сравнивали поведение кошек на протяжении 30 мин до и 30 мин после проигрывания сигналов тревоги местных обезьян тити Callicebus toppini и саки Pithecia rylandsi или других громких и продолжительных вокализаций данных видов, использованных для контроля: первое заставляло их прекращать прежнюю активность и уходить прочь из этого района. И наоборот: обработка контрольными вокализациями не влияла на поведение кошек: они продолжали находиться в засаде, отдыхать или двигаться в прежнем направлении. Расстояние, на которое отходит оцелот при проигрывании сигналов тревоги, также значимо выше, чем проходимое после проигрывания контрольных звуков. Данные непосредственных наблюдений за поведением оцелотов, обнаруженных и окрикиваемых обезьянами vs оставшихся незамеченными, подтверждает результаты экспериментов (Adams, Kitchen, 2018).
Иными словами, обнаруженный хищник страдает не меньше жертвы, знающей что на неё могут охотиться.
Функция сдерживания хищника у сигналов тревоги султанок
J. Woodland, Zarinah Jaafar and Mary-Lou Knight
За небольшим исключением (Perrins 1968; Smythe 1970), этологи всегда считали, явно или по умолчанию, что функция сигналов тревоги — предупреждать особей своего вида о присутствии хищника. Части из них казалось самоочевидным, что сигнализирующие особи тем самым рискуют быть им схваченными, снижая свои шансы на выживание (Brereton 1959; Maynard Smith 1965; Hirth and McCullogh 1977). Именно эволюционные следствия этого второго предположения [как выяснилось дальше, неверного, по крайней мере, в таком общем виде. Здесь и далее прим.публикатора] вызвали наибольший интерес к этой проблеме [окрикивание хищника это «альтруизм» или нет?]. Если бы удалось показать, что такое поведение не только альтруистично, но его наличие и действие характеризует какие-то социальные группы, не являющиеся группами родственников [а таковы очень многие социумы; у ряда птиц и млекопитающих дисперсия молодых и «притяжение» вселенцев организованы так, что они направленно подбираются из наименее родственных особей.], это дало бы доказательства «группового отбора», ахиллесовой пяты социобиологических взглядов на популяционную саморегуляцию (Wynne-Edwards 1962). Противники последнего могут сослаться на «бритву Оккама» и утверждать, что никакой это не альтруизм, а самосохранение, пока не доказано обратного (Смайт, 1970; Чарнов и Кребс, 1975). [И будут правы!]
Однако мы не уверены, что эти авторы учли все возможные объяснения функции сигнала тревоги [а правило Оккама требует не умножать сущностей/не усложнять объяснения сверх необходимости, т. е. сигнальных эффектов предъявления демонстраций сигналов, которые были надежно показаны в полевых наблюдениях или лабораторных опытах]. Наши наблюдения за сигналом «тревоги» восточной, или черноспинной, султанской курочки (Porphyrio porphyrio L.) указывают на ещё одну возможность, упущенную в рассуждениях выше — такого рода сигналы адресованы хищнику, а не сородичам [или не только сородичам, их издают также для сдерживания хищника в рамках «психологической дуэли».]. Здесь мы развиваем идею Смайта (Smythe, 1970), предположившего, что белое пятно, как бы вспыхивающее при поднятии хвоста косуль, оленей и других крупных копытных (обычно в момент убегания от какой-то опасности) у различных травоядных млекопитающих «вызывает преследование» со стороны хищника [видимо, действуя как запускающий охоту релизер]. Однако наша интерпретация реакций хищника на сигналы тревоги иная; данные по султанкам доказывают что он сдерживает хищника [блокируя переход к охоте, а то и заставляя уйти].

Черноспинная султанка
Материал и методы
Черноспинная султанка (сегодня рассматривается как отдельный вид Porphyrio melanotus) крупная, иссиня-чёрная пастушковая птица с ярко-белыми кроющими подхвостья [склонная при движении или на месте «кланяться», резко подымая хвост вверх, когда белое пятно «вспыхивает» также, как у косуль). Местом исследования стали 70 га лагуны на северо-востоке Нового Южного Уэльса, окружённой разорванным кольцом ситниковых зарослей, дорогами, за ними — небольшим по протяжённости пастбищем; в самой лагуне было много зарослей Eleocharis sphacelata, высокой (1-2 м) травы из сем.Осоковых.
Большую часть времени султанки проводили, рассеявшись поодиночке среди тростника, но также отваживались выходить на берег для пастьбы (ощипывания растительности). Они предпочитали окраины некоторых улучшенных пастбищ примерно в 30-50 метрах от кромки воды. Там иногда они объединяются в открытые группы (непостоянного состава). При дистанции между птицами не более 3 м наблюдалась определённая степень координации между членами стаи. Размер группы изменчив, с модальным классом от 4 до 8 птиц, вполне обычны и одиночки.
Наиболее обычными хищниками, представляющими опасность для курочек, здесь были рыжая лисица, домашняя собака и человек. Птицы относятся к людям весьма настороженно, и авторы полагали что реакция на подходящих людей может моделировать реакцию на скрадывающего их наземного хищника. Подходы «потенциальной опасности» и фиксация реагирования птиц на неё, составившие данные табл.1, производились одними и теми же людьми за 3 мес. до начала репродуктивного сезона
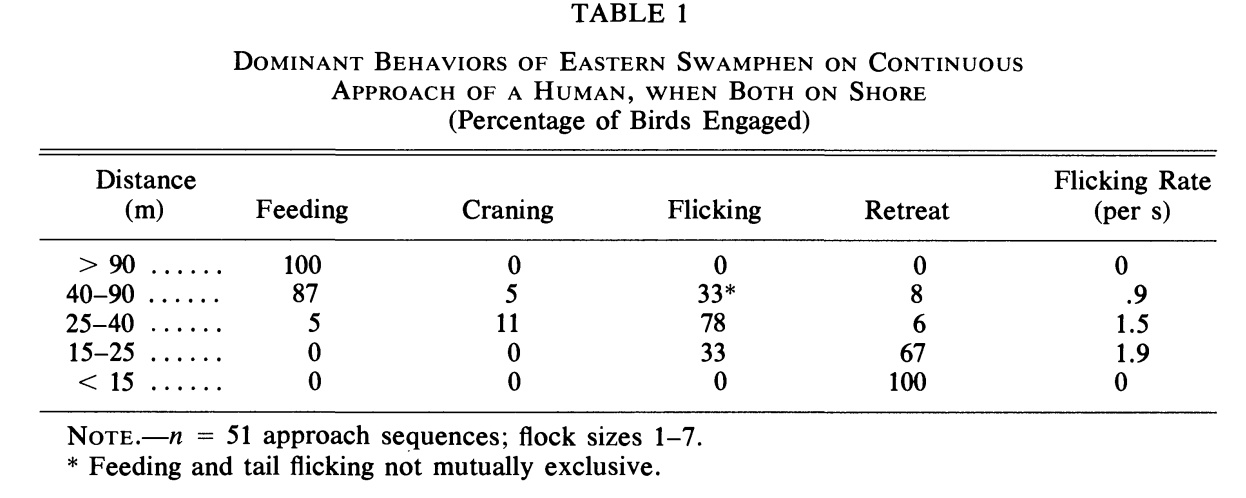
таблица 1. Преобладающее поведение в группах султанок на разной дистанции подхода к ним человека, % вовлечённых птиц. Столбцы слева направо: классы расстояния, м; кормление; выпрямление и вытягивание шеи; «вздёргивания» хвостом; отступление; частота «вздёргиваний», раз/с
* Кормление и вздёргивания хвоста могут совмещаться друг с другом.
Для сбора данных один и тот же человек («тестировщик») шел медленным шагом по тропинке примерно в 10 м от кромки воды к султанкам, кормящимся на открытом пастбище. Реакцию птиц на него, и дистанции, на которых наступают различные реакции, фиксировали наблюдатели, укрывшиеся на вышке. Некоторые дополнительные замечания обсуждаются ниже. Несколько таких подходов записаны на видеопленку.
Результаты
На открытой местности типичный ответ султанок на подход «тестировщика» был следующим. На дистанции около 90 м птицы впервые проявили тревогу, взглянули на подходящего, выпрямившись и вытянув шею, что ненадолго прервало кормление. При более близком подходе они начали «мелькать» белым подхвостьем: резко поднимали и опускали хвост (как бы вздёргивая его: все действие занимало менее 2 секунд), почему «вспыхивало» белое пятно, направляемое в сторону незваного гостя. Взмахивающие хвостом птицы одновременно от=ходят на пару шагов от наблюдателя; взмахивания повторяются неоднократно, их частота зависит от расстояния между птицей и нарушителем (табл.1).
«Вспыхивающее» пятно очень сильно привлекает внимание, даже когда дёргающая хвостом султанка находится на периферии поля зрения «тестировщика». При этом кормление продолжается. На данной дистанции птицы были повернуты более или менее в сторону от приближающегося человека (но под прямым углом к их возможной линии отступления), они одновременно кормятся и вздёргивают хвостом.
Когда нарушитель приблизился к птицам метров на 40, большинство из них перестали кормиться и отступили в лагуну, скорость отхода зависела от расстояния, которое надо было пройти. Вздёргивания хвоста продолжались, завися от скорости. У медленно идущих птиц они были нерегулярными, спазматическими, идущие быстрей каждый шаг сопровождали взмахом хвоста; бегущие птицы не дёргали им вовсе. На этом этапе интенсивность «вспыхивания» белых подхвостий, направленного к источнику опасности, конечно, уменьшилась, но сама демонстрация была различимой. Остановившись у кромки воды, султанки продолжили дёргать хвостом, пока «тестировщик» не сблизился с ними на ~ 20 м, затем слетели к зарослям E.sphacelata в лагуне.
Перед интерпретацией этих данных приведём наблюдения, дающие дополнительную информацию, важную для их понимания, подкрепляющую рабочую гипотезу о «сдерживании потенциальной опасности». В ряде подходов «тестировщик» имитировал преследование стай из 4-5 птиц на дистанции ~40 м, когда они пробуют отступать в лагуну. При таких обстоятельствах [т. е. отступив] султанки успокаиваются, возвращаясь к кормлению. При этом птицы, ближайшие к подходящему, продолжают настойчиво «дёргать» хвостом, в то время как более дальние практически перестают это делать. В 1-минутных наблюдениях за 10 стаями средняя скорость вздёргивания хвоста у 16 «ближайших» султанок была 36,4 взмаха/мин (с пределами 4-60 м), тогда как у 31 наиболее «дальних» (находившихся на 3-6 м далее от наблюдателя по сравнению с первыми) — только 2,6 (с диапазоном 0-14 м). «Ближайшие» особи при вздёргивании хвоста всегда поворачивали его к «опасности», ориентация более дальних при исполнении тех жен демонстраций была случайной.
В зарослях E.sphacelata султанки рассредотачиваются гораздо сильней, обычны дистанции от 10 до 20 м между особями в одной обособленной куртине среди воды. При приближении к ним здесь на лодке птицы, не могущие немедленно скрыться в зарослях, просто отворачивались от неё и вздёргивали хвостом, но при наличии рядом убежища птицы сразу скрывались там без демонстраций. (То же самое делали кормящиеся на берегу среди зарослей ситника у края лагуны: в ответ на приближение пригибались и прятались, не демонстрируя). В этих условиях демонстрации начинались лишь при сближении с «опасностью» на 20 м: птицы выпрямлялись и вздёргивали хвост, ориентируя «вспыхивающее» пятно в сторону нарушителя или под углом к линии между ними, но не более 450.
Эти весьма заметные «вспышки» часто — первый признак присутствия птиц. Нарушитель на лодке мог обогнуть отдельные кущи E.spacelata по кругу. При этом кормящиеся внутри султанки оставались практически на одном месте; держались всё время выпрямившись [маркёр побуждений страха и бегства] и оглядываясь назад, но при вздёргивании хвоста каждый раз поворачивали тело так, чтобы пятно направлялось на нарушителя. Частота вздёргиваний с приближением «опасности» здесь также росла (до 2 в с).
Обсуждение
Из данных наблюдений прямо следует, что вздёргивания хвоста у султанок не только адресуются возможному хищнику, но и сигнализируют ему о бдительности данной особи, её готовности к бегству [чем сдерживаются готовящееся нападение]. Во-первых, на пастбищах не выявлено различий между демонстрациями одиночек и птиц в группах. Если [вслед за социобиологами] принять, что демонстратор рискует, тревожная сигнализация у одиночек была бы исключена отбором — риск есть, а выигрыш отсутствует. Однако одиночки вовсю использовали вздёргивания, и часто при обстоятельствах, когда сигнал виден только потенциальному хищнику. И наоборот, при возможности сразу скрыться в убежище, птицы делали это, а не демонстрировали [не пробовали «предупредить сородичей», хотя риск минимален]. И только когда укрытие было слишком далеко или невелико, не скроешься, птицы активно демонстрировали в сторону потенциальной опасности.
И наоборот, птицы, у которых было доступное укрытие, предпочли спрятаться вместо того, чтобы пытаться «предупредить сородичей». Только когда их прикрытие, скорее всего, оказывалось бесполезным, они раскрывали себя и щелкали хвостом.
Во-вторых, если сигналы тревоги рискованны, непонятно их учащение с приближением к ним «тестировщика»: в этом случае риск сильно увеличивается именно для данной особи, но вовсе необязательно для всех остальных. В рамках теории отбора родичей можно рассуждать, что подобное учащение вздёргиваний хвоста увеличивает число родственников, которые может быть будут предупреждены о растущей опасности [здесь сразу вспомнился мем «хайли лайкли»]. Возможно, однако, иное объяснение [исходящее из проксимальных, а не ультимативных причин, что разумней делать этологам]: если сигнал [будучи социальным релизером] адресуется хищнику, то его приближение увеличивает стимуляцию, усиливает в султанках состояние страха и стресса, и они демонстрируют эти релизеры интенсивней, чтобы потенциальную опасность всё же остановить, побудить хищника отказаться от атаки [обычный для этологов механизм релизерного действия демонстраций: проблема — демонстрация — усугубление проблемы — более эффективная демонстрация или более интенсивное демонстрирование прежней и пр., пока проблемная ситуация не будет успешно разрешена или птица не проиграет взаимодействие, скажем, её таки схватят]. Кроме того, в рамках обоих названных [социобиологических] объяснений совершенно неясно, почему в преследуемой стае демонстрируют самые уязвимые для хищника птицы, почему они делают это максимально открыто и, главное, в направлении грозящей опасности (и тем более точно «в направлении», чем она ближе)?
В-третьих, вздёргивание хвоста как визуальный сигнал сравнительно малоэффективное средство оповещения сородичей. В части случаев при кормлении в группах султанки действительно оповещаются об опасности, наблюдая за товарками, обнаружившими «тестировщика», видя, как они смотрят на него, выпрямившись и вытянув шею, или же дёргают хвостом. В других случаях это не получалось, и «тестировщик» зашёл на пастбище значительно дальше дистанции, с которой султанки отступают к краю лагуны, прежде чем его обнаружили. В этом случае курочки, оставшиеся позади, просто бежали к лагуне без каких-либо демонстраций. Если принять [точку зрения социобиологов], что сигналы тревоги рискованны для демонстратора, но выигрышны для воспринимающих особей, намного лучшим средством оповещения об опасности были бы акустические сигналы. Их могут услышать султанки, не видящие демонстратора, они много хуже локализуются, что уменьшает шанс хищника определить, кто именно подал сигнал. На деле всё прямо наоборот: несмотря на открытую местность, демонстраторы снова и снова «рекламируют» себя хищнику: для сородичей вздёргивание хвостом это скрытый сигналом, а вот для возможного хищника очень даже явный.
Резюмируя: поведение демонстратора здесь скорее эгоистичное, а не альтруистическое, поскольку оно наиболее точно идентифицирует опасность, и «указывает» её степень, но доступность сигнала сородичам уменьшается.

Тасманийская зеленоногая камышница
Наблюдения Ридпата (1972) за родственной султанкам тасманийской зеленоногой камышницей Tribonyx mortierii дали дополнительные данные в поддержку рабочей гипотезы авторов. Она внешне сходна с султанкой, близка по размеру, но нелетающая и не имеет белого подхвостья. Демонстрация вздёргивания хвоста в присутствии хищника у неё есть, но с другим визуальным образом [адресуемым хищнику]: крылья быстро выдвинуты в горизонтальное положение, потом столь же резко убираются. Она исполняется почти исключительно в ответ на присутствие клинохвостого орла Aquila audax. Авторы считают, что самое важное в этом сигнале в отличие от вздёргивания хвоста у султанок — резкое изменение силуэта камышницы в горизонтальной плоскости (а не в вертикальной), что увеличивает заметность сигнала сверху, то есть воздушным хищником.
До сих пор обсуждался в основном адресат сигналов тревоги. В согласии со Smythe, 1970 авторы доказывают, что это в первую очередь хищник, а не сородичи. Однако они расходятся со Smythe, 1970, 1977 в интерпретации эффекта подобных сигналов — это не провокация атаки или преследования хищником (в том числе преждевременной и полезной для жертвы), а его остановка, [«давление» на него релизерами], чтобы отказался от атаки и искал себе других жертв [менее осторожных и поэтому не демонстрирующих]. Smythe (1970, 1977) считал, что жертвы будут вести себя так, как он предположил, потому они действуют в рамках ограниченного бюджета времени. Однако на деле бюджет времени с большей вероятностью будет ограничен у хищника, а не у жертвы, особенно если последняя травоядна, как это обычно бывает [и каковы виды в данной работе].
Стабильная коэволюция хищника и его жертвы [своего рода «гонка вооружений», соревнование брони и снаряда] продолжается лишь пока жертва остается в ней «на шаг впереди» хищника [«лиса бежит за своим обедом, но заяц — за своей жизнью»]. По крайней мере, многие хищные позвоночные [даже самые специализированные] значительные трудности в обнаружении и поимке жертв, почему вынуждены преодолевать значительные расстояния между успешными охотами, охотиться только в определенное время суток, собирать падаль, оппортунистически изменять свой рацион [массовыми легкодоступными кормами, от ягод и мелких зверьков до пищевых отходов или насекомых, несмотря на свою специализацию], охотиться на наиболее уязвимых жертв (как молодые, старые, больные или раненые). Подтверждающие данные предоставлены Rudebeck (1950, 1951) для пернатых хищников-орнитофагов, Schaller (1967) для тигров Panthera tigris и оленей Axis axis, Cervus duvauceli, Cervus unicolor, Mech (1966, р.51) для волков Canis lupus и лосей Alces alces (в той же работе он рассматривает и другие такие исследования). Более того, в данной литературе дважды упомянута реакция хищника, предсказываемая в рамках гипотезы сдерживания тревожными сигналами. Elliott et al. (1977) выявил, что африканский лев Panthera leo не преследует добычу, знающую о его местонахождении. Здесь особенно уместна цитата из Schaller (1967, с. 288):
«Тревожный лай оленя, направленный на затаившегося тигра, явно достаточный признак для кошки, что дальнейшее преследование бесполезно, поскольку затем она обычно поднимается из своего укрытия и уходит».
Из приведенных выше исследований мы пришли к выводу, что у видов, для которых обычно обсуждаются сигналы тревоги (хищники и жертвы из числа высших позвоночных), первые часто испытывают большие трудности при поимке добычи. Жертвы не только кажутся обладающими сенсорным преимуществом, позволяющим обыкновенно обнаруживать хищника до того, как он сможет успешно атаковать, но и бдительное животное, находящееся в расцвете сил, по-видимому, имеет чрезвычайно высокую вероятность избежать поимки. Как следствие, время охоты для хищника очень «дорого» и должно использоваться оптимально. Поэтому когда жертва специфическим сигналом тревоги «сообщает» хищнику, что он обнаружен, а оно информировано об опасности, отслеживает её динамику, то хищнику выгоднее всего убраться прочь и искать других.
[С точки зрения итоговой приспособленности] было бы крайне полезно, если б сигналы тревоги жертв одновременно рекламировали бы их бдительность и физическую форму [тем более что это связанные вещи]. Для волка вид и/или скорость убегающего лося достаточны для решения, стоит ли преследовать (Mech, 1966, р.121). «Свечи» (подпрыгивания высоко вверх у многих копытных, в том бегущих прочь от опасности или беспокойства, самый яркий пример — сайгаки Saiga tatarica) — ещё лучшее средство достичь этой цели ловким способом достижения той же цели. «Свечи» отмечены у многих животных с белыми пятнами на крупе, их функции часто описывается как усиление данного сигнала тревоги (Guthrie 1971). Однако Shaller (1967) зафиксировал аналогичное поведение у гаура Bos gaurus, у которого нет пятна на крупе. Поэтому авторы предполагают, что «свечи» — сигнал ad hoc [движения намерения, ещё не прошедшие ритуализацию], адресованный хищнику и останавливающий его (здоровое животное прыгает бодро, энергично и часто, имолодое, раненое или больное — увы, первое удержит от нападения). И наоборот, в демонстрациях, провоцирующих хищника на нападение, с последующим обманом (отводящие демонстрации у птиц), визуальный образ построен вокруг изображения травмы, вроде «сломанного крыла» или «хромания», которое оказывается ложным, как только привлечённый подобным хищник будет отвлечён хищника достаточно далеко от гнезда и птенцов (например, Hinde 1952, стр. 148).
Charnov и Krebs (1975) объясняют происхождение и последующую эволюцию сигналов тревоги исходя из того, что они адресованы хищнику и рискованны для демонстратора [это неединичный пример очччень цитируемой статьи с выводами, не соответствующими фактическим данным]. Однако [отчасти противореча самим себе ] они предположили, что реакция сородичей на сигналы тревоги может быть увеличивать приспособленность самого демонстратора [что верно для каждого члена группы]. Если выигрыш перевешивает риск, сигналы тревоги будут прогрессивно развиваться на основе индивидуального отбора. Скажем, система работала бы, если б сигналы тревоги собирали рассеянных птиц в стаю, что давало бы демонстратору больший выигрыш, чем его сородичам (скажем он оказывался бы в наиболее защищённом центре стаи, чем компенсировал бы риски сигнализации). Замечательная теория, но нужная ли?
Мы бы согласились с тем, что сигналы тревоги могут быть перехвачены хуже информированными сородичами и информация может быть использована в их интересах. Кроме того, демонстратор может еще больше снизить свои шансы на поимку, действуя согласованно с другими особями в стае или, что еще лучше, используя наилучшее знание об опасности, её приближении и пр., чтобы занять еще более выгодное положение в стае сравнительно с сородичами. Однако всё сказанное — отдельные оборонительных реакций при появлении хищника, тогда как Чарнова и Кребса рассматривают их в едином комплексе, их объяснение требует, чтобы весь комплекс реакций развивался одновременно. Все приведённые наблюдения за султанками резко расходятся и с тем и с другим.
Список литературы
Brereton, J. Le G. 1959. Evolved regulatory mechanisms of population control. Pages 81-93 in G.W. Leeper, ed. The evolution of living organisms. Melbourne University Press, Melbourne.
Charnov, E. L., and J. R. Krebs. 1975. The evolution of alarm calls: altruism or manipulation? Am. Nat. 109:107-112.
Elliott, J. P., I. McTaggart Cowan, and C. S. Holling. 1977. Prey capture by the African lion. Can. J.Zool. 55:1811-1828.
Guthrie, R. D. 1971. A new theory of mammalian rump patch evolution. Behaviour 38:132-145.
Hinde, R. A. 1952. The behaviour of the great tit (Parus major) and some other related species. Behaviour Suppl. 2.
Hirth, D. H., and D. R. McCullough. 1977. Evolution of alarm signals in ungulates with special reference to white-tailed deer. Am. Nat. 111:31-42.
Maynard Smith, J. 1965. The evolution of alarm calls. Am. Nat. 99:59-63.
Mech, L. D. 1966. The wolves of Isle Royale. U.S. Natl. Park Serv. Fauna Natl. Parks US, Fauna Ser. 7.
Perrins, C. 1968. The purpose of the high-intensity alarm call in small passerines. Ibis 110:200-201.
Ridpath, M. G. 1972. The Tasmanian native hen, Tribonyx mortierii. I. Patterns of behaviour. CSIRO Wildl. Res. 17:1-51.
Rudebeck, G. 1950. The choice of prey and modes of hunting of predatory birds with special reference to their selective effect. I. Oikos 2:65-88
Rudebeck, G., 1951. The choice of prey and modes of hunting of predatory birds with special reference to their selective effect. II. Oikos 3:200-231.
Schaller, G. B. 1967. The deer and the tiger. University of Chicago Press, Chicago.
Smythe, N. 1970. On the existence of «pursuit invitation» signals in mammals. Am. Nat. 104:491-494.
Smythe, N. 1977. The function of mammalian alarm advertising: social signals or pursuit invitation? Am. Nat. 111:191-194.
Wynne-Edwards, V. C. 1962. Animal dispersion in relation to social behaviour. Oliver & Boyd, Edinburgh.
Рекомендуем прочесть
 Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции
Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции Групповые адаптации без группового отбора?
Групповые адаптации без группового отбора? Манипуляционно-исследовательская активность у морских птиц: поведенческая реакция на предъявленные на колонии искусственные объекты
Манипуляционно-исследовательская активность у морских птиц: поведенческая реакция на предъявленные на колонии искусственные объекты Номогенетическое
Номогенетическое Разное про запаховую коммуникацию
Разное про запаховую коммуникацию Ссылки по биологии
Ссылки по биологии Взлёт и падение универсальных «базовых эмоций» Экмана
Взлёт и падение универсальных «базовых эмоций» Экмана Зачем большим пёстрым дятлам всю зиму охранять территории?
Зачем большим пёстрым дятлам всю зиму охранять территории?
