

Безбивневые слонихи в Горонгосе
Резюме. Понимание (микро)эволюционных следствий антропогенной эксплуатации биоресурсов делается все важнее по мере увеличения доли изъятия особей охотниками и рыболовами из популяций опромышляемых видов. Авторы изучили влияние браконьерской добычи слоновой кости во время войны в Мозамбике (1977-1992) на эволюцию африканских саванных слонов (Loxodonta africana) в национальном парке Горонгоса (Gorongosa). Браконьерство вызвало сильный отбор, благоприятствовавший отсутствию бивней у самок на фоне быстрого сокращения популяции.
Её обследование выявило паттерны наследования бивней, соответствующие действию доминантного гена, связанного с Х-хромосомой и летального для самцов. Полногеномный анализ показал существование всего двух кандидатных генов [участвующих в развитии безбивневости], AMELX и MEP1a. Вполне известно их участие в развитии зуба млекопитающих, включая формирование эмали, дентина, цементного вещества зубов и пародонта. Один из них (AMELX) у людей ответственен за развитие Х-сцепленного доминантного синдрома, у женщин уменьшившего рост боковых резцов верхней челюсти (гомологичных бивням слона). У мужчин поражения значительно более тяжёлые, хотя и несмертельные.
Это исследование предоставляет доказательства быстрого действия отбора, опосредованного браконьерством, обеспечившего потерю ключевой анатомической характеристики у целевого вида. [Отбор в пользу самок оказывается губительным для самцов, т.е. он сексуально-антагонистичен. Утопающий хватается и за соломинку: так в условиях браконьерства очень вредные генные варианты стали спасением. Последнее получилось именно потому, что у этого полигинного вида самцы в отличие от самок не очень важны для социальной организации, группы разного уровня возглавляются старшими самками и объединяются в (более рыхлые) группы более высокого уровня за счёт персонализованных дружественных взаимоотношений между ними. Будь социальная жизнь этого вида иной, отбор в пользу безбивневости мог снизить жизнеспособность популяций вместо наблюдаемого повышения. Прим.публикатора].
Избирательное уничтожение видов, чьи анатомические особенности (бивни, рога, и пр.) определяют их трофейную и товарную ценность, питает незаконную торговлю дикими животными (1) с многомиллиардными оборотами. Она прямо угрожает выживанию экологически важной мегафауны во всем мире (2, 3).
Крупнейшие травоядные особенно уязвимы к перепромыслу из-за необходимости больших территорий для существования популяций, требовательности к местообитаниям, небольшой численности и медленного размножения (длительный оборот генераций) (4, 5). Эти виды — «экосистемные инженеры»: своей деятельностью они преобразуют среду обитания, создают мозаику специфических нарушений в определённом масштабе пространства, и этим регулируют экологические процессы (5-8). Поэтому антропогенный отбор фенотипов, влияющих на это поведение, запустит каскадные изменения в функционировании соответствующих экосистем. Однако большинство исследований отбора, производимого человеческой деятельностью, выполнено на мелких видах, где легче изучать эволюционные изменения (9, 10). Как происходят фенотипические изменения, обусловленные промыслом, у крупнейших наземных животных в мире, до сих пор остается неясным (в какой степени они происходят? с какой скоростью? посредством каких механизмов?).
Все войны в Африке усиливают эксплуатацию запасов диких животных и сокращают их численность (11). Военные действия долгое время тесно переплетались с торговлей слоновой костью (12-14). За годы мозамбикской гражданской войны на территории национального парка Горонгоса (1977-1992 гг.) население крупных травоядных сократилось более чем на 90% (15), обе стороны конфликта охотились на слонов ради бивней (15, 16). [Это была не гражданская война, а агрессия расистских режимов ЮАР и Северной Родезии против народа Мозамбика, избравшего социализм, для этого они использовали прокси из местных правых и трайбалистов. Без внешней агрессии, бывшей частью “холодной войны” Запада против СССР и его союзников, а также народов, привлечëнных советским опытом, эта война бы даже не началась. Прим.публикатора]. Однако интенсивное браконьерство в Африке увеличило случаи появления слонов без бивней, почти всегда (или исключительно) у самок (таблица S3). В парке Горонгоса безбивневые слоны-самцы не отмечены (таблица S2). Анализ видеоматериалов прошлых лет и данных сегодняшних наблюдений (дополнительные материалы) показывает, что резкое сокращение популяции слонов Горонгосы сопровождалось почти трехкратным учащением встреч самок без бивней, с 18,5% (n = 52) до 50,9% (n = 108) (двухвыборочный тест на равенство пропорций с коррекцией по непрерывности, Р < 0,001) (рис.1А).

Леса Горонгосы
Чтобы проверить, было ли учащение безбивневости слоних случайным событием, связанным с серьезным сокращением популяции [проход через «бутылочное горлышко» с соответствующими изменениями генофонда] (17), авторы смоделировали наблюдаемое сокращение численности в Горонгосе с 1972 года (n = 2542 особи) по 2000 год (n = 242) (15), положив в основание 0-гипотезы равные вероятности выживания слоних с бивнями и без (см. Методы). Моделирование показало исключительно низкую вероятность что учащение безбивневости произошло без участия отбора (гипергеометрическое распределение, P = 1,8 * 10-15) (рис.1B). За эти 28 лет относительная выживаемость слоних без бивней была более чем в пять раз выше, чем с бивнями (оценка максимального правдоподобия = 5,13, 95% доверительный интервал 3,98-6,60) (рис.1С). Можно заключить, что «бутылочное горлышко» популяции слонов в Горонгосе сопровождалось сильным отбором в пользу фенотипа без бивней.
При наличии сильного отбора против слонов с бивнями, мы увидим его «геномную подпись»: дивергентные изменения генома, наблюдаемые по мере того как в популяции фенотип без бивней теснит фенотип с бивнями. Авторы секвенировали полные геномы 18 слоних из образцов крови (7 с бивнями, 11 без). Эти генетические тексты авторы сопоставили с последовательностями аннотированного генома саванновых африканских слонов (Loxafr3.0) и сгенерировали выравнивания с покрытием ~30х для 13 образцов и покрытием 14 для 5 образцов (дополнительные материалы). Используя выборки с покрытием 30× (6 с бивнями, 7 без), авторы вычислили дистанцию Таджимы D (18) для разных участков генома в неперекрывающихся окнах размером 10 кб. [Дистанция Таджимы — мера различий между двумя образцами генетического разнообразия: отражает среднее количество попарных различий и количество участков сегрегации, каждое из которых масштабируется таким образом, что ожидается, что они будут одинаковыми в нейтрально развивающейся популяции постоянного размера. Прим.публикатора]. В геномах обеих групп слоних выявлен небольшой избыток редких вариантов, ответственный за отрицательные значения D (без бивней: -0,27, с бивнями: -0,2).
Однако геномы слоних с бивнями включали значимо меньше редких вариантов, чем без них (двухвыборочный t-критерий Уэлча: P<0,0001) (рис.1D и дополнительные материалы), что согласуется с большим падением численности первых.
Чтобы оценить эволюционную реакцию на отбор, авторы определили частоты разных фенотипов у взрослых самок, родившихся после войны, с 1995 г. по 2004 г.
В потомстве выживших самок доля безбивневости (33%, n = 91) значимо выросла сравнительно с таковой до конфликта (18,5%, двухвыборочный тест на равенство пропорций с поправкой на непрерывность, P = 0,046) (рис.1A) и превзошла ожидаемую в отсутствие отбора (гипергеометрическое распределение, P = 4,3*10-8) (рис.1B). Эти результаты указывают на генетическую детерминированность отсутствия бивней и эволюционную реакцию слонов Горонгосы на отбор, вызванный браконьерством. Учитывая доказательства наследуемости отсутствия бивней и его специфичность для слоних Горонгосы, авторы посчитали, что данный признак детерминирован локусом, сцепленным с полом (17, 19-21). Поэтому они искали закономерность наследования, которая объяснить наблюдаемый паттерн различий. Фенотипы, присутствующие только у самок, обычно приписываются доминантному гену в Х-хромосоме, летальному для самцов (22). Соответственно, авторы сопоставляли фенотипы матерей и потомков в Горонгосе, чтобы проверить рабочую гипотезу, что безбивневость наследуется моногенно, доминантным геном, сцепленным с Х- и летальным для самцов. По этой гипотезе, самки с бивнями несут 2 аллеля дикого типа (X+X+), самцы один (X+ Y). Тогда слонихи с бивнями рожают дочерей только с бивнями; самки без бивней — гетерозиготы по аллелю безбивневости (X+X−), унаследовавшие аллель дикого типа от отца, безбивневые самцы нежизнеспособны. Отсюда, дочери безбивневых слоних будут 2х типов, с бивнями или без, в пропорции 1:1, и жизнеспособна лишь половина самцов, родившихся от гетерозиготных матерей. Следовательно, 2/3 потомства самок без бивней также будет слонихами (в предположении полной пенетрантности фенотипа Х-).
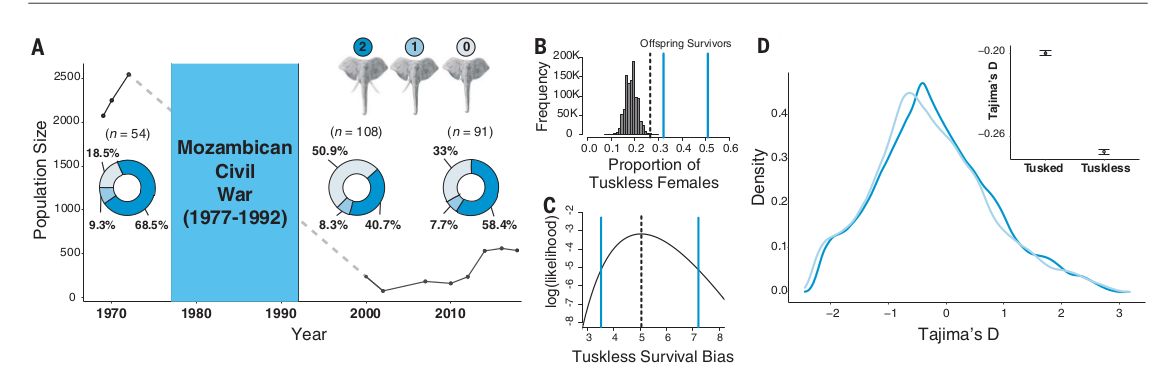 Рисунок 1. Демографические изменения слоновьей популяции нацпарка Горонгоса за годы гражданской войны и интервенции в Мозамбике, свидетельства отбора в пользу безбивневости. А. Изменение численности популяции (линия) и долей среди слоних безбивневых, одно- и двубивневых (серо-голубой, голубой и синий сектора круговой диаграммы) до войны (n=54), выживших после неё (n=108), .и в родившемся у них 1м поколении (n=91). Линия — минимум численности по данным авиаучëтов (30); пунктир — нет данных. Доли приведены по современным обзорам и историческим фотографиям, не включены особи, у которых нельзя было определить, есть бивни и сколько их.
Рисунок 1. Демографические изменения слоновьей популяции нацпарка Горонгоса за годы гражданской войны и интервенции в Мозамбике, свидетельства отбора в пользу безбивневости. А. Изменение численности популяции (линия) и долей среди слоних безбивневых, одно- и двубивневых (серо-голубой, голубой и синий сектора круговой диаграммы) до войны (n=54), выживших после неё (n=108), .и в родившемся у них 1м поколении (n=91). Линия — минимум численности по данным авиаучëтов (30); пунктир — нет данных. Доли приведены по современным обзорам и историческим фотографиям, не включены особи, у которых нельзя было определить, есть бивни и сколько их.
B. Моделирование по распределению Валлениуса доли безбивневых слоних, ожидаемой после «бутылочного горлышка» популяции, в предположении отсутствие отбора по данному признаку. Сплошные вертикальные линии — наблюдаемый % таких слоних среди выживших в годы войны и родившихся сразу после (1995 — 2004 гг.). Пунктир — верхний перцентиль моделируемого распределения.
C. Вероятность большего шанса на выживание самок без бивней в Горонгосе (ордината) в зависимости от эмпирического соотношения частот выживших слоних без бивней к таковым с бивнями (логарифмированная, абсцисса).
Пунктир — оценка максимального правдоподобия; сплошные синие линии — её 95% доверительный интервал. D. График плотности общегеномных дистанций D Таджимы для слоних с 2мя бивнями (темно-синий) и без бивней (светло-голубой), рассчитанный в окнах размером 10 кб. Врезка. Среднее значение ± 1 SE общегеномного D для каждого из 3х вариантов наличия бивней.
У слоних с 2мя бивнями 91,3% дочерей также имели 2 бивня (n = 21 с двумя бивнями, 1 без бивня, 1 с одним бивнем) (рис.2А). У безбивневых слоних, наоборот, родилось примерно поровну дочерей с бивнями или без [n = 19 (40,9%) и n=21 (44,7%) соответственно, тест на равенство пропорций в двух выборках с поправкой на непрерывность, P = 0,42] (рис. 2A). У матерей обеих фенотипов в небольшом % случаев рождались «промежуточные» дочери с одним бивнем [у 2хбивневых одна (4,3%), у безбивневых семь (14,9%), тест на равенство пропорций P = 0,37]. У потомства безбивневых слоних также значимо повышена доля самок (учитывались те, кому ≤5 лет, n = 67), до 65,7%. Это значимо отличается от 0-гипотезы о равном соотношении полов (точный биномиальный тест, P = 0,027), и неотличимо от доли в 2/3 самок, ожидаемой при полной летальности безбивневых самцов (Р = 0,90) (рис.2В).
Напротив, у слоних с бивнями не выявлено значимых отклонений соотношения полов в потомстве от 1:1 (самок 54,2%, что значимо не отличается от 50%, n = 48, P = 0,67) (рис.2B). Предшествующие исследования африканских слонов также не выявили отклонений от 1:1 при рождении детëнышей (23). Тогда можно заключить, что отмеченный сдвиг этого соотношения к 2/3 коррелирует с выраженностью безбивневости.
В целом, 87,1% ассоциаций между фенотипами матери и потомства соответствуют однолокусной доминантной, X — сцепленной модели наследования. Сдвиг соотношения полов, связанный с безбивневостью, не более чем на 1% отличался от ожидаемого при полной смертности самцов. Необъяснëнная изменчивость признака, включая одностороннее отсутствие бивней (коли оно генетически детерминировано), [можно объяснить, предположив] эпистатические взаимодействия между локусами (минимум двумя) влияющие на внешнее выражение морфологии бивня. Известно разнообразие связей генотип-фенотип, влияющих на изменение морфогенеза зубов, весьма разнообразны, их отличительные мутации [в смысле таких изменений] идентифицировались как популяционно- или даже семейно-специфичные (24).
Кроме того, эпигенетическое формирование паттернов [строения зуба] и мозаичная инактивация Х-хромосомы могут вести к изменчивой реализации в фенотипе одних и тех же мутаций, связанных с неразвитием зубов, даже между однояйцевыми близнецами (25). Тем не менее данные из Горонгосы подтверлили рабочую гипотезу о детерминированности безбивневого фенотипа как минимум одним Х-сцепленным доминантным локусом, летальным для самцов. Он даёт максимальный эффект; возможно, есть также локусы-модификаторы, дополнительно влияющие на экспрессию признака.
Авторы идентифицировали этот локус, использовав полногеномные данные; в разных участках генома они искали «подписи» селективных процессов, связанных с недавним интенсивным браконьерством. Они анализировали геномы для выявления наиболее вероятных локусы-кандидатов на роль «мишеней» отбора по признаку сцепленному с полом и летальному для самцов. Сперва они просканировали весь геном на предмет локусов, показывающих сильный недавний отбор, специфичный для фенотипа без бивней, и сосредоточились на геномах безбивневых особей (n = 11).
Хотя этот признак в Горонгосе не зафиксировался (рис.1), при анализе геномов безбивневых слоних авторы ждали сильных сигналов недавнего отбора.
Использовав метод скользящего окна из окон размером 10 кб с шагом 2 кб, они отыскали 2 «подписи» селективных давлений, ожидаемых сообразно гипотезе отбора по Х-сцепленному доминантному признаку, летальному для самцов. Секвенировав геномы 13 слоних с покрытием 30× (6 с бивнями, 7 без), авторы сперва поискали области генома, демонстрирующие избыточную гетерозиготность по аллелям, специфическим для безбивневого фенотипа, гетерозиготность определяли количественно по отклонению от равновесия Харди-Вайнберга. Дальше искали локусы, отличающиеся неравновесием по сцеплению (LD), они содержат мутации высококоррелированные между собой именно у безбивневых фенотипов, среди образцов без клыков. LD определяли количественно с помощью векторов к однонуклеотидным полиморфизмам (SNP) слоновьих геномов (mLD). Сродство их к разным участкам подсчитывается в программном обеспечении RAiSD (26). Далее авторы сравнивали все геномы самок с бивнями (n = 7) vs без них (n = 11) для поиска участков генетических «текстов», на которых отмечена наибольшая дивергенция между ними, её сила и значимость оценивается Fst.
При верности рабочей гипотезы ожидается, что в геномах слоних с бивнями нет специфической мутации или мутаций, напрямую определяющих отсутствие бивней. Они же с куда меньшей вероятностью имеют в геномах SNP, сцепленные с такого рода мутацией (-ями) [и передающиеся эффектом попутного транспорта. Прим.публикатора]. Мы количественно определили генетическую дифференциацию, используя обе метрики нормализованных различий по аллельным частотам, Fst (27), и среднее число попарно взятых различий [между участками], Dxy (28). Участки генома, дающие значимый выброс относительно «фона» (превышение значимо большее 95% квантиля общегеномного распределения) по всем четырем использованным показателям (гетерозиготность, LD, Fst и Dxy) считались возможной «мишенью» отбора на безбивневость (гены-кандидаты полезного признака). Перекрывание выбросов по данным показателям выявило 305 таких кандидатных участков генома (рис.2C).
Если отсутствие бивней доминантный и Х-сцепленный признак, определяющий его ген или гены также должен быть там. Среди 305 возможных участков авторы отфильтровали свойственные Х-хромосоме. Это всего 8 отрезков ДНК, попадающих в два смежных участка генома. 5 из них перекрывают область ~100 кб, содержащую X-связанный ген изоформы амелогенина (AMELX) (рис.2D). Он кодирует белок внеклеточного матрикса, участвующий в биоминерализации эмали, и предположительно регулирует как образование периодонта, так и работу генов, связанных с образованием цемента (29, 30). Несколько мутаций в пределах этого локуса связаны с недоминерализацией эмали и хрупкостью зубов у людей (30).
Делеция в синтенной области Х-хромосомы человека (Xp22.2), охватывающая AMELX и несколько смежных генов, ведёт к несовершенному амелогенезу, сопровождающемуся Х-сцепленным доминантным синдромом, смертельным для мужчин (31). У женщин она вызывает ряд аномалий зубного ряда, включая микродонтность и/или неразвитие боковых резцов верхней челюсти (31), гомологов бивням слона. Примечательно, что искаженная инактивация Х-хромосомы при несовершенном амелогенезе способствует выраженной фенотипической изменчивости у гетерозигот (32). Предыдущие исследования показали, что самцы генноинженерных мышей, «сконструированных» с делецией в этой области, могут выжить при принудительной экспрессии гена синтетазы голоцитохрома с-типа человека (HCCS), расположенном по соседству с AMELX (33). Высокий LD, отмеченный на данном участке Х-хромосомы у безбивневых слоних (рис.2D и рис. S1) рождает предположение, что пространственная близость между AMELX и соседними локусами с самцовой летальностью обеспечила связь отсутствия бивней и самцовой летальности в Горонгосе.
Остальные три отрезка-кандидата на Х-хромосоме охватывают необработанный псевдоген FAM115B и прилежающую межгенную область. Авторы не нашли какой-либо известной связи между FAM115B и одонтогенезом или развитием челюстно-лицевой области.
Учитывая потенциальное влияние эпистаза на экспрессию морфологии бивня в [популяции] Горонгосы, авторы дополнительно просканировали весь геном в поиске областей, наиболее дивергировавших именно у безбивневых слоних сравнительно с имеющими бивни (об этом судили по Fst или Dxy, P < 0,001) (рис.3A) или имеющих низкое генетическое разнообразие, характерное для безбивневого фенотипа (относительное разнообразие = p безбивневых слонов − pслонов с бивнями, P < 0,001. Этим критериям удовлетворили 3 смежных геномных участка (значимые Fst и относительное разнообразие, при P < 0,001) (рис.3B). Эта область перекрывается с геном МЕР1а, отдельным аутосомным геном на первой хромосоме. Он кодирует альфа-субъединицу меприна, матриксного металлопротеина, играюшего важную роль в минерализации дентина путем его синтеза из предшественника, сиалофосфопротеина дентина (DSPP). Аномалии структуры DSPP связаны с несколькими нарушениями развития зуба, включая дисплазию дентина, к ведущую к аномалиям формирования корней зубов и преждевременной их потере (34).
Мыши c неработающим геном MEP1a (MEP1a−/−) показывают значительные изменения минеральной плотности костной ткани дентина (35). Найденные авторами три «окна» входят в более протяжённый геномный интервал (~6 Мб), характеризующийся увеличенным LD (P < 0,001) (рис.3C и рис. S2), что говорит о недавнем положительном отборе на этом участке. Оба гена, AMELX и MEP1a функционально ассоциированы с развитием некоторых областей зуба млекопитающих, включая эмаль, дентин, цемент и периодонт (рис.4). Однако анализ дивергентных полиморфизмов и структурных вариантов генома (включая делеции, дупликации и варианты числа копий) между [особями разных] морфам бивней ни для одного локуса не выявил очевидных причинно-следственных связей между соответствующим генным вариантом и искомым признаком (дополнительные материалы).
Таким образом, безбивневость, опосредованная браконьерством во время войны в Мозамбике, на уровне генов обусловлена недавним отбором как минимум по одному Х-сцепленному (AMELX) и одному аутосомному локусу (MEP1a). Непосредственная физическая связь гена AMELX с ближайшими к нему летальными для самцов локусами на Х-хромосоме, такими как HCC (31, 33), может обеспечивать наблюдаемую картину наследования безбивневости в Горного сельского как доминантного признака, летального для самцов и сцепленного с полом.
При верности данной интерпретации в данном исследовании обнаружен редкий случай производимого человеком сексуально-антагонистического отбора, благоприятствующего признаку самок, несмотря на его [дотоле неизвестную] вредность самцам. Учитывая временные рамки отбора, скорость эволюционной реакции и известное наличие выбранного фенотипа до события отбора, отбор постоянной генетической вариации в этих локусах является наиболее правдоподобным объяснением быстрого роста случаев отсутствия бивней в течение этих 15 лет войны.
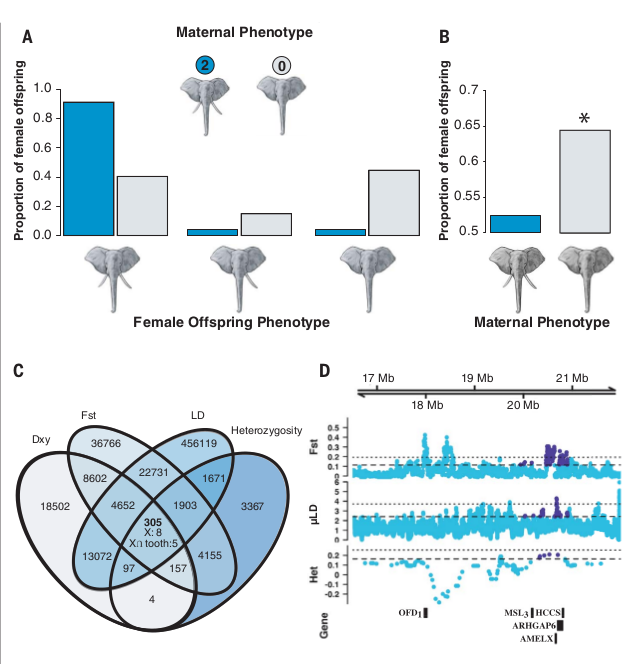 Рисунок 2. Доказательства Х-сцепленного доминантного наследования [безбивневости] с мужской летальностью, основанного на действии выявленных кандидатных локусов. A и B. Выводы о закономерностях наследования исходя из характера потомства в популяции. А. Распределение альтернативных фенотипов у дочерей слоних, имеющих бивни (темно-синий) и без бивней (светло-голубой). B. Наблюдаемое соотношение полов в потомстве самок с бивнями и без. Звездочка — значимое отклонение от 1:1 у матерей без бивней (биномиальный тест, Р = 0,027), но не у матерей с бивнями (Р = 0,67). C и D. Доказательства отбора по локусу-кандидату, сцепленному с полом. C. Диаграмма Венна из 4 метрик генетической дифференциации между участками генома слоних с бивнями и без бивней, вычисленная методом скользящих окон. Диаграмма показывает число перекрывающихся окон в 5% хвостах распределения каждой статистики (искомые кандидаты в области пересечения всех 4х).
Рисунок 2. Доказательства Х-сцепленного доминантного наследования [безбивневости] с мужской летальностью, основанного на действии выявленных кандидатных локусов. A и B. Выводы о закономерностях наследования исходя из характера потомства в популяции. А. Распределение альтернативных фенотипов у дочерей слоних, имеющих бивни (темно-синий) и без бивней (светло-голубой). B. Наблюдаемое соотношение полов в потомстве самок с бивнями и без. Звездочка — значимое отклонение от 1:1 у матерей без бивней (биномиальный тест, Р = 0,027), но не у матерей с бивнями (Р = 0,67). C и D. Доказательства отбора по локусу-кандидату, сцепленному с полом. C. Диаграмма Венна из 4 метрик генетической дифференциации между участками генома слоних с бивнями и без бивней, вычисленная методом скользящих окон. Диаграмма показывает число перекрывающихся окон в 5% хвостах распределения каждой статистики (искомые кандидаты в области пересечения всех 4х).
В число этих статистик входят 2 меры генетической дифференциации между фенотипами с бивнями и без бивней (Fst и Dxy), а также LD и отклонение гетерозиготности от равновесия Харди-Вайнберга в выборке безбивневых особей. Цифры в пересечении диаграммы — число соответствующих окон на Х-хромосоме и то подмножество окон, которое перекрывает известные гены развития зубов (X ∩ зуб). D. Увеличенные манхэттенские графики Fst, LD и гетерозиготности (Het) показывают расположение в геноме пяти окон, перекрывающих гены развития зубов: AMELX с фланкирующими областями. Ось х графика — геномные координаты, пунктир — верхние 5% и 1% квантили доверительных интервалов для выбросов показывающих значимые различия метрик. Темно-синие точки значимые выбросы (Р < 0,05) в пределах кандидатного отрезка гена, светло-голубые — отрезки генетического «текста» без значимых выбросов. Область-кандидат содержит 23 гена, но для ясности помечены только гены, которые, как известно, участвуют в развитии зубов и/или мужской смертности.
 Рисунок 3. Аутосомный локус-кандидат на детерминацию безбивневости у слонов в Горонгосе. A-C. Скаффолд 0 в хромосоме 1 африканского слона, [на основании разных метрик дистанций между фенотипами] выделяющий локус-кандидат на [детерминацию] безбивневости, MEP1a. Ось Х везде — хромосомные координаты, показаны изменения этих метрик вдоль хромосомы (голубой) , выделены участки генома со значимыми отличиями по ним (тёмно-синий). A. Генетическая дивергенция (Fst) между морфами с бивнями и без. B. Относительное разнообразие внутри генома у безбивневых особей (Dπ = πбезбивневых слонов − πслонов с бивнями). Низкие значения показывают участки генома с низким относительным генетическим разнообразием у безбивневых особей C. Неравновесие по сцеплению, количественно определяемое с использованием SNP-векторов (MLD) среди безбивневой выборки. Крайние отклонения в каждом распределении (Р < 0,001) отмечены темно-синими точками. Светло-синие точки указывают на неэкстремальные значения. Сплошные вертикальные линии показывают положение MEP1a в хромосоме 1 генома слона. Пунктир — границы прилежащей области с высокой плотностью повышенного LD.
Рисунок 3. Аутосомный локус-кандидат на детерминацию безбивневости у слонов в Горонгосе. A-C. Скаффолд 0 в хромосоме 1 африканского слона, [на основании разных метрик дистанций между фенотипами] выделяющий локус-кандидат на [детерминацию] безбивневости, MEP1a. Ось Х везде — хромосомные координаты, показаны изменения этих метрик вдоль хромосомы (голубой) , выделены участки генома со значимыми отличиями по ним (тёмно-синий). A. Генетическая дивергенция (Fst) между морфами с бивнями и без. B. Относительное разнообразие внутри генома у безбивневых особей (Dπ = πбезбивневых слонов − πслонов с бивнями). Низкие значения показывают участки генома с низким относительным генетическим разнообразием у безбивневых особей C. Неравновесие по сцеплению, количественно определяемое с использованием SNP-векторов (MLD) среди безбивневой выборки. Крайние отклонения в каждом распределении (Р < 0,001) отмечены темно-синими точками. Светло-синие точки указывают на неэкстремальные значения. Сплошные вертикальные линии показывают положение MEP1a в хромосоме 1 генома слона. Пунктир — границы прилежащей области с высокой плотностью повышенного LD.
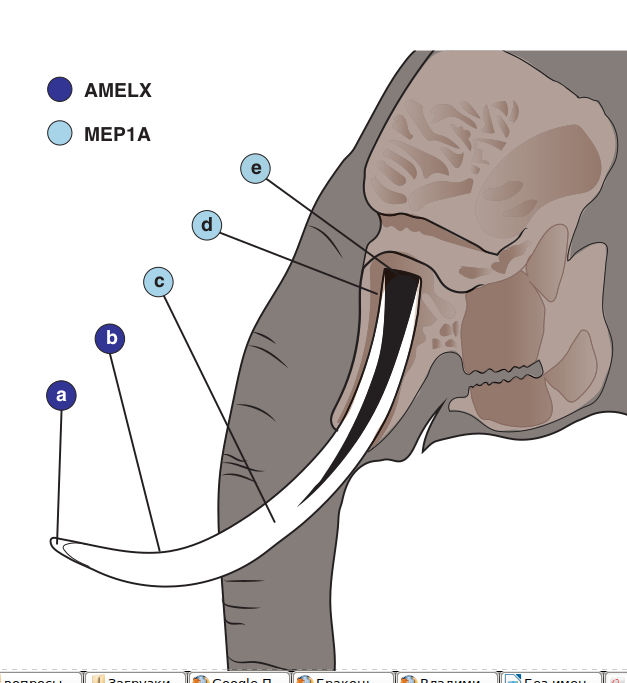
Рисунок 4. Возможное действие локусов-кандидатов на морфологию бивня. Поперечный разрез бивня [т.е. резца] африканского слона показывает расположение (а) эмали, (б) цемента, (в) дентина (слоновая кость), (г) периодонта и (е) корня зуба. Темно-синим обведены области, подверженные влиянию гена-кандидата AMELX (точно известному или предположительному), светло-синим круги — влиянию MEP1a. Ни один из генов, фигурировавших в анализе, не влияет на формирование зубной пульпы (черное на поперечном сечении бивня).
[Необходимые пояснения биоинформатических терминов. сейчас прочитывают относительно короткие участки (риды), а потом с помощью программ собирают их в единый геном. Покрытие — это среднее число ридов на участок. Естественно, где-то больше, где-то меньше, где-то разрешить последовательность трудно из-за повторов. Поэтому 14 — это неплохое покрытие, 30 — очень хорошее (как у авторов статьи), 5 — весьма посредственное. Длина рида зависит от использованного метода секвенирования. Скаффолд — это этап сборки, длинная последовательность, которая ещё не с хромосому, но в которую собрано много коротких прочтений. Дистанция Таджимы показывает степень не-нейтральности эволюции. Прим.публикатора]
Однако точные генетические и развитийные механизмы и механизмы, ведущие к безбивневости и/или нежизнеспособности самцов, пока неизвестны. Хотя самцы без бивней не отмечены ни в Горонгосе, ни при обследованиях сколько-нибудь значительного числа особей в наиболее интенсивно изучаемых популяций африканских слонов (17, 21, 36, 37), имеются неподтвержденные сообщения о самцах без клыков в нескольких местах (20, 38, 39). Нам неизвестны исследования, которые твердо установили бы,что встречаемость самцов без бивней превышает правдоподобно объяснимую редкими травмами или ошибкой наблюдателя (дополнительный текст и таблица S3), однако нельзя исключить возможность [действия] альтернативных генетических механизмов и/или появления безбивневости при взаимодействии генотипа с определёнными внешними условиями. Более того, промежуточные фенотипы с одним бивнем обычно встречаются в семейных группах, также включающих безбивневых самок (17, 20, 37).
Хотя данные по Горонгосе согласуются с Х-сцепленным доминантным признаком, приводящим к летальному исходу у самцов, общеконтинентальные паттерны степени выраженности и наследуемости бивней могут определяться не обнаруженным отбором в его пользу, а другими факторами. Например, географическими различиями по LD между AMELX и прилежащими локусами [определяющими] летальность самцов, дополнительными локусами в других частях видового генома, индивидуальными вариациями в паттернах активации Х-хромосомы или каким-то совсем другим механизмом. Необходимы дальнейшие исследования, чтобы определить точное число и местонахождение генных вариантов, кодирующих безбивневость; потребуются сравнительное изучение нескольких популяций, чтобы выявить географическую структуру изменчивости и характер наследования [по этим генам], обеспечивающий детерминацию данного признака.
[Последствия] социальных конфликтов и коммерческого промысла могут переплестись, подрывая популяции эксплуатируемых видов (11, 40). Однако большинство известных случаев эволюционных изменений, вызванных перепромыслом, происходят постепенно в течение более длительных периодов времени, и специфические эффекты именно промысла может быть трудно отделить от других факторов (9, 41-44). Данное исследование показывает, как внезапный всплеск беспорядков [в дотоле спокойной стране] может вызвать резкие и стойкие эволюционные сдвиги у долгоживущих животных даже в условиях экстремального сокращения популяции. В Горонгосе и сами слоны, и [обычное ранее] присутствие у них бивней крайне значимы для восстановления экосистемы. Слоновьи бивни — это многоцелевые инструменты, которые используются для добычи подземных продуктов питания и минералов (45, 46), а также для выдалбливания и отслаивания коры, что может вызывать гибель деревьев (47, 48). Преобразуя среду обитания и создавая мозаику нарушений, слоны выступают экосистемными инженерами. Рост численности слонов может катализировать переход от лесов к пастбищам в больших масштабах (45); отсутствие у них бивней может нарушить или значительно изменить все названные процессы, дестабилизировать природные сообщества вместо прежнего поддержания.
Рекомендуем прочесть
 Быстрая эволюция холодостойкости у анолисов в южном Техасе
Быстрая эволюция холодостойкости у анолисов в южном Техасе Быстрая эволюция в связи с урбанизацией: примеры
Быстрая эволюция в связи с урбанизацией: примеры Мини-рецензия на книгу Менно Схилтхёйзена «Дарвин в городе. Как эволюция продолжается в городских джунглях»
Мини-рецензия на книгу Менно Схилтхёйзена «Дарвин в городе. Как эволюция продолжается в городских джунглях» Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов
Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов Экспериментальная эволюция гуппи в речках Тринидада.
Экспериментальная эволюция гуппи в речках Тринидада. Эволюция в пространстве возможностей: Дарвин и Вавилов
Эволюция в пространстве возможностей: Дарвин и Вавилов Урбанизация и антропогенез: неожиданные параллели
Урбанизация и антропогенез: неожиданные параллели Теории биологической эволюции с позиций современного развития системной биологии
Теории биологической эволюции с позиций современного развития системной биологии
