![Print PDF Данные об изменениях глобальной температуры (микроклимат) получены с метеостанций, меряющих ее на высоте 1-1, 2 м над уровнем высокой травы. Температура, значимая для организмов, связана с микроклиматом, и […]](http://www.socialcompas.com/wp-content/uploads/2023/07/why-are-forests-turnin-465x190.jpg "Глобальное потепление изменяет леса лишь по нарушениям лесного полога")
 Данные об изменениях глобальной температуры (микроклимат) получены с метеостанций, меряющих ее на высоте 1-1, 2 м над уровнем высокой травы. Температура, значимая для организмов, связана с микроклиматом, и почти всегда отличается. Рельеф и растительность, лесная особенно, создают собственную температуру и более или менее устойчиво поддерживают её даже вопреки планетарным изменениям. Это ведёт к рассогласованию изменений растительности и планетарной температуры. Предполагается, что по мере её повышения разные виды растений в сообществе будут всё больше сменяться более теплолюбивыми, это так называемая термофилизация.
Данные об изменениях глобальной температуры (микроклимат) получены с метеостанций, меряющих ее на высоте 1-1, 2 м над уровнем высокой травы. Температура, значимая для организмов, связана с микроклиматом, и почти всегда отличается. Рельеф и растительность, лесная особенно, создают собственную температуру и более или менее устойчиво поддерживают её даже вопреки планетарным изменениям. Это ведёт к рассогласованию изменений растительности и планетарной температуры. Предполагается, что по мере её повышения разные виды растений в сообществе будут всё больше сменяться более теплолюбивыми, это так называемая термофилизация.
Однако подобная смена зависит от микро-, не макроклимата: если сообщество демонстрирует буферные свойства, то изменения первого отстают от изменений второго, и с ними запаздывает смена растительности — тем больше, чем буферные свойства сильнее выражены. Как известно, последнее имеет место в лесах, исследованных в разных районах Европы Zellweger et al. (2020). Отставание смены видов растений и животных, обеспеченное устойчивостью микроклимата вопреки происходящему потеплению, получилo название климатический долг (climatic debt).
Трёхмерная структура лесного полога обеспечивает максимальное постоянство условий существования сравнительно с другими сообществами, благодаря затенению, перемешиванию воздуха и испарительному охлаждению.
Он защищает кустарники, травянистый ярус и подстилку от экстремальной жары, и эта буферизация динамично меняется вслед за изменением мозаично-оконной структуры сообщества, включая старение деревьев 1го яруса, созданные им изменения кроны, с последуюшим выпадением и зарастанием «окна». Логично предположить: микроклимат сообщества и, соответственно, сохраняемость его видов в условиях потепления (или его противоположность, термофилизация) прямо зависят от развития нарушений, создающих » прорехи» полога или их противоположности, уплотнения верхнего яруса с возрастом в охраняемых местообитаниях.
[Для изучения буферной роли лесов] авторы сочетали измерения температуры под пологом леса в 100 массивах зоны умеренных лесов Европы, с анализом изменений растительности на 2955 участках из 56 регионов, где она постоянно сохранялась, а каждый участок повторно обследовали с перерывом от 12 до 66 лет (рис.1А и рис. 2B). Связав эмпирические данные о температуре in situ (микроклимат) с состоянием полога в континентальном масштабе, авторы прогнозируют изменения температуры подлеска за вегетационный период, основываясь на взаимосвязи между пологом и буферизацией температур макроклимата (21) (рис.1BC).
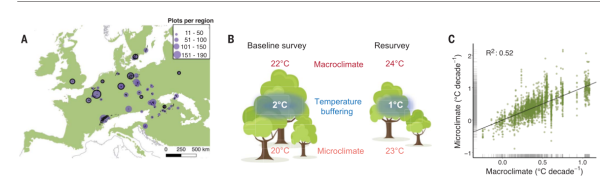 Рисунок 1. При трансформации полога лесного массива микроклимат меняется много сильней, чем общее потепление за тот же период. (A) Расположение 2955 переобследованных лесных массивов, принадлежащих биому лесов умеренной зоны (черные точки) в 56 регионах Европы (размер фиолетовых точек отражает объем выборки, легендированный вверху справа). Авторы собрали репрезентативные данные об изменении микроклимата в 100 лесных насаждениях, по 10 в каждом из 10 регионов (черные круги; эффективное n = 96), чтобы определить максимум их буферных свойств на протяжении вегетационного периода в зависимости от состояния полога (23) (относительно изменений макроклимата рис. S2). (B) Примерная схема определения изменения микроклимата в нижних ярусах лесного массива. Слева сообщество при базовом, справа при повторном обследовании. Здесь полога стал менее сомкнутым (или сперва был более сомкнутым, чем потом), почему буферизация лесом общего потепления уменьшилась с 2 до 1°C. Вследствие этого микроклимат сообщества теплеет быстрее (с 20 до 23°C, внизу картинки) чем макроклимат (от 22 до 24°C, вверху картинки). Взаимосвязь между изменением лесного полога и буферизацией максимальных температур макроклимата оценивали эмпирически для всего района исследований (рис. S2) (21). (C) Зависимость скорости изменения микроклимата конкретных лесных сообществ (точки) от скорости изменений макроклимата по всей данной территории (то и другое в градусах за декаду). Черная разделяющая линия представляет соотношение 1:1 (точное следование первого за вторым). И микро- и макроклимат стали намного теплее, см. текст, конкретные измерения скоростей изменения того и другого даны серой штриховкой по осям. Темпы изменения первого на 45% изменчивей темпов изменения второго, последние объясняют лишь около половины вариаций первых (предельная (условная) доля объясненной дисперсии R2, равное 0,52 (0,69). Все статистические результаты основаны на моделях со смешанными эффектами с регионом в качестве свободного члена (случайного эффекта регрессии).
Рисунок 1. При трансформации полога лесного массива микроклимат меняется много сильней, чем общее потепление за тот же период. (A) Расположение 2955 переобследованных лесных массивов, принадлежащих биому лесов умеренной зоны (черные точки) в 56 регионах Европы (размер фиолетовых точек отражает объем выборки, легендированный вверху справа). Авторы собрали репрезентативные данные об изменении микроклимата в 100 лесных насаждениях, по 10 в каждом из 10 регионов (черные круги; эффективное n = 96), чтобы определить максимум их буферных свойств на протяжении вегетационного периода в зависимости от состояния полога (23) (относительно изменений макроклимата рис. S2). (B) Примерная схема определения изменения микроклимата в нижних ярусах лесного массива. Слева сообщество при базовом, справа при повторном обследовании. Здесь полога стал менее сомкнутым (или сперва был более сомкнутым, чем потом), почему буферизация лесом общего потепления уменьшилась с 2 до 1°C. Вследствие этого микроклимат сообщества теплеет быстрее (с 20 до 23°C, внизу картинки) чем макроклимат (от 22 до 24°C, вверху картинки). Взаимосвязь между изменением лесного полога и буферизацией максимальных температур макроклимата оценивали эмпирически для всего района исследований (рис. S2) (21). (C) Зависимость скорости изменения микроклимата конкретных лесных сообществ (точки) от скорости изменений макроклимата по всей данной территории (то и другое в градусах за декаду). Черная разделяющая линия представляет соотношение 1:1 (точное следование первого за вторым). И микро- и макроклимат стали намного теплее, см. текст, конкретные измерения скоростей изменения того и другого даны серой штриховкой по осям. Темпы изменения первого на 45% изменчивей темпов изменения второго, последние объясняют лишь около половины вариаций первых (предельная (условная) доля объясненной дисперсии R2, равное 0,52 (0,69). Все статистические результаты основаны на моделях со смешанными эффектами с регионом в качестве свободного члена (случайного эффекта регрессии).
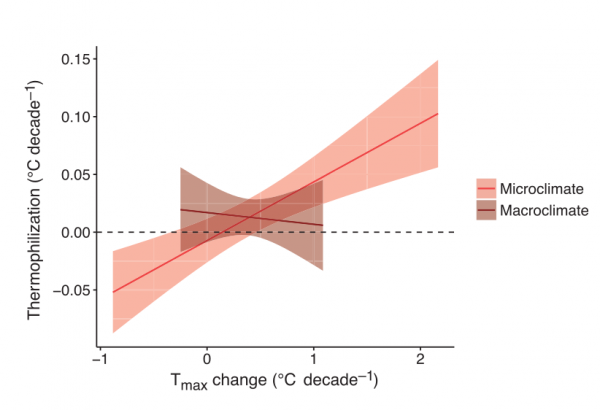 Рисунок 2. Термофилизация в растительных сообществах лесного подлеска связана с изменением микро-, а не макроклимата. Термофилизация фитоценозов (ордината) ускоряется с усилением потепления микроклимата до температурного максимума за вегетационный период (Tmax). Ось Х — темпы изменения Тmax внутри лесных сообществ (микро-) и метеостанциями (макроклимат). Связь термофилизации с темпами потепления макроклимата статистически не значима, см. текст. Для обоих зависимостей показана линия регрессии ± 95% доверительный интервал.
Рисунок 2. Термофилизация в растительных сообществах лесного подлеска связана с изменением микро-, а не макроклимата. Термофилизация фитоценозов (ордината) ускоряется с усилением потепления микроклимата до температурного максимума за вегетационный период (Tmax). Ось Х — темпы изменения Тmax внутри лесных сообществ (микро-) и метеостанциями (макроклимат). Связь термофилизации с темпами потепления макроклимата статистически не значима, см. текст. Для обоих зависимостей показана линия регрессии ± 95% доверительный интервал.
Изменения полога во времени сильно варьировали в 56 регионах, в диапазоне от -110% (появление множества прорех) до +113% (сильное уплотнение полога) (1-й и 99-й процентили распределения соответственно). При этом изменение полога в среднем по выборке значимо не отличалось от 0 (+2,6%; модели со смешанными эффектами P = 0,426; рис. S3). Прогноз изменений температуры подлеска за время между базовым и повторным обследованием каждого участка производили при помощи мы ранее опубликованной статистической модели, оценивающей изменения приземной температуры в зависимости от сомкнутости кронового яруса (21, 23) (рис. S2).
Согласно ему, максимальные температуры в последние десятилетия существенно выросли, в среднем (±SEM) показателями 0,40 ± 0,04 и 0,38 ± 0,03°C за десятилетие для потепления микро- и макроклимата соответственно (обе оценки темпов потепления основаны на смешанных данных).модели эффектов: Р < 0,001). Микроклимат, однако, был на 45% изменчивей (1-й и 99-й процентили: от -0,32 до 1,36°C за десятилетие), макро- (1-й и 99-й процентили: от -0,08 до 1,08°C за десятилетие) (рис. S4). Связь между обоими изменениями была высоко значимой (Р < 0,001), однако 48% общего изменения микроклимата (наклон: 1,05, R2= 0,52, Р < 0,001) (рис.1С) были оставались необъяснимы исходя из изменений макроклимата. Отсюда естественно предположить, что первый не зависит от общего потепления напрямую, но формируется каждым отдельным лесным сообществом, и зависит от его состояния, в первую очередь от сохранности полога. Чем она лучше выражена, тем сильней буферные свойства, ослабляющие связь между параметрами и наоборот, чем больше «прорех», тем непосредственней потепление формирует микроклимат конкретного массива.
Для количественной оценки термофилизации, авторы определили температурные оптимумы и пределы температурной толерантности всех видов сосудистых растений, присутствующих в данных [т.е. произрастающих на обследованных площадках] в зоне их нынешнего распространения. Используя данные о предпочитаемой температуре для каждого конкретного вида, авторы для каждого обследованного сообщества рассчитали скорость изменения максимально допустимых температур за время между базовым и повторным обследованием (14, 23) (рис. S6).
Мы ожидали, что изменения максимальных значений сродства к температуре будут наиболее тесно связаны с изменениями максимальных температур микро- и макроклимата в течение вегетационного периода (23). Биотическая реконструкция температурной динамики, построенная на изменениях видового состава сообществ, широко использовалась для оценки последствий климатических изменения климата для сообществ, сохраняющих те или иные наземные и морские таксоны (12-14, 16). Итоговые темпы термофилизации на 2955 постоянных участках варьировали от -0,84 до 1,05°C за десятилетие, со средним значением (±SEM) 0,01 ± 0,01°C [что значимо не отличается от нуля (P=0,09) (23)].
Темпы термофилизации растительности подлеска положительно связаны со скоростью изменений микро-, но не макроклимата была положительно связана со скоростью потепления микроклимата [шкалирования оценка наклона регрессии для зависимости от микроклимата 0,02, 95-й доверительный интервал (ДИ): от 0,01 до 0,03, Р < 0,001, то же для макроклимата: -0,002, ДИ: от -0,01 до 0,01, Р = 0,70) (рис.2).
Для количественной оценки влияния микроклимата на наблюдаемый климатический долг каждого из исследованных лесных сообществ, авторы из скорости изменения микроклимата (то и другое в единицу времени, DTmicro/dt) — вычли темпы изменения растительности DTplant/Dt). Это же вычитание скоростей сделали и для изменения макроклимата (DTmacro/Dt) – (DTplant/Dt)] для каждого из фокальных графиков. Средние значения (±SEM) микро- и макролиматического долга почти совпали (0,38±0,04°C/10 лет vs 0,37±0,04°C/10 лет), однако климатические долги, рассчитанные с использованием данных макроклимата, недостаточно отражают вариабельность микроклиматического долга (рис. S7).
Наибольшее потепление микроклимата отмечено в массивах с деградацией полога, сократившей буферные способности сообщества. Это были участки, где исходно микроклиматический долг был наибольшим (рис.3А) [и быстро понизился по мере развития прорех]. Местный нагрев, вызванный деградацией полога, снижает способность растительных сообществ нижних ярусов приспосабливаться к быстрому потеплению макроклимата. еагировать на такие высокие темпы потепления. Напротив, климатический долг понижен в массивах с уплотнением полога. Усиление буферных свойств сообщества вызвало охлаждающий эффект в течение всего вегетационного периода.
Эти зависимостм остаются невидимыми, если климатический долг рассчитывать лишь по данным о макроклимате (рис.3B). Поэтому реалистические оценки изменений лесов под прессом глобального потепления, требуют данных о долговременных изменениях микроклимата. [Или, что то же самое, структуры лесного полога, определяющего буферные свойства сообщества].
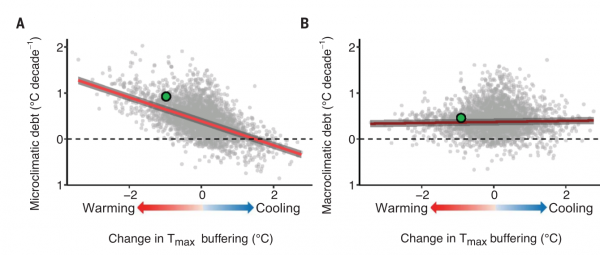 Рисунок 3. Буферизация температуры под пологом леса, если он сохранился, объясняет климатический долг лесных биогеоценозов. (А) Климатический долг, рассчитанный на основе изменения температуры микроклимата (т.е. микроклиматический долг), увеличивается по мере того как слабеет буферизация (Tmax) при деградации лесного полога [наклон: -1,88, предельный (условный) R2= 0,51 (0,75), P < 0,001]. Ось Х — изменения лесного полога, отрицательные — распространение в нём «прорех» и нагрев нижних слоёв сообщества (и меньшая буферизация Tmax); положительные — охлаждение нижних слёв биогеоценоза при уплотнении полога и большей буферизации (Tmax). (B) Климатические долги, рассчитанные по скоростям изменения макроклимата (т.е. долг макроклимата) почти что не связаны с разницей в буферизации [наклон: 0,13, предельный (условный) R2= 0 (0,25), P = 0,06]. Показана линия регрессии ± 95% доверительный интервал (серые полосы). Зеленые точки — примерный микро- и макроклимат после изменения (уменьшения) буферизации Tmax, показанного на рис.1B.
Рисунок 3. Буферизация температуры под пологом леса, если он сохранился, объясняет климатический долг лесных биогеоценозов. (А) Климатический долг, рассчитанный на основе изменения температуры микроклимата (т.е. микроклиматический долг), увеличивается по мере того как слабеет буферизация (Tmax) при деградации лесного полога [наклон: -1,88, предельный (условный) R2= 0,51 (0,75), P < 0,001]. Ось Х — изменения лесного полога, отрицательные — распространение в нём «прорех» и нагрев нижних слоёв сообщества (и меньшая буферизация Tmax); положительные — охлаждение нижних слёв биогеоценоза при уплотнении полога и большей буферизации (Tmax). (B) Климатические долги, рассчитанные по скоростям изменения макроклимата (т.е. долг макроклимата) почти что не связаны с разницей в буферизации [наклон: 0,13, предельный (условный) R2= 0 (0,25), P = 0,06]. Показана линия регрессии ± 95% доверительный интервал (серые полосы). Зеленые точки — примерный микро- и макроклимат после изменения (уменьшения) буферизации Tmax, показанного на рис.1B.
Динамика структуры полога запускает изменения микроклимата внутри лесных массивов, сильно отличающиеся от внешних по отношению к ним изменений макроклимата [и даже противоположны им]. Это существенно для прогноза реакции биоразнообразия на изменения климата и землепользования (например, лесопользования). Взаимодействуя между собой, они стимулируют появление новых термальных сред [внутри сообщества, и только потом самое потепление начнёт действовать].
Ожидаемо усиление аномальной жары (4), при котором многие виды и сообщества сильно пострадают при наличии в пологе прорех из-за удаления старых деревьев или их отмирания (24). Возникающие последствия крайне серьезны: поскольку леса сохраняют большую часть биоразнообразия суши, а многие экосистемные услуги и средства к существованию критически зависят от биоразнообразия лесов (1, 25). Поэтому управляющим лесами и директивным органам в условиях потепления следует не столько обращать внимание на само потепление, сколько учитывать влияние разных методов лесопользования на [структуру сообщества, его буферные свойства и тем самым на] микроклимат в усилиях по сохранению биоразнообразия лесов.
Полученный результаты подтверждает гипотезу, что темпы термофилизации растительности лесного массива определяются прежде всего скоростью изменения микроклимата под пологом [и заданы сдвигом структуры этого полога за соответствующий период (10, 12)], а не скоростью изменения макроклимата [иными словами, не планетарным потеплением как таковым]. Это прямое эмпирическое доказательство определяющего [в конечном счёте] влияния изменений микроклимата на ответ организма на климатические изменения, что часто игнорируется при использовании макроклиматических данных для изучения подобных ответов (8, 26, 27).
Рост климатического долга природных сообществ в условиях потепления означает, что всё больше видов попало и вынуждено выживать в пессимальных климатических условиях, что потенциально ускоряет утрату биоразнообразия. Эта работа показывает, что изменения структуры сообщества, [ослабляющие или усиливающие его буферные свойства, действуя через изменения микроклимата] могут как усиливать действие планетарного потепления, так и защищать сообщество от него. [Фактически потепление может менять состав растительности только в нарушенных сообществах и в первую очередь это происходит по местам нарушений]. Поэтому климатические долги, рассчитанные по данным макроклимата (13, 20), следует интерпретировать с осторожностью, фактически их следует пересмотреть с учётом нарушенности сообществ.
Рекомендуем прочесть
 Глобальное потепление, опустынивание/деградация почв и засухи в аридных регионах
Глобальное потепление, опустынивание/деградация почв и засухи в аридных регионах Лес и климат. Влияние изменений климата на леса России: зафиксированные воздействия
Лес и климат. Влияние изменений климата на леса России: зафиксированные воздействия Изменения климата: факты, модели и механизмы
Изменения климата: факты, модели и механизмы Сумма про антропогенные изменения климата — 2
Сумма про антропогенные изменения климата — 2 Как изменение климата способствует быстрой эволюции
Как изменение климата способствует быстрой эволюции Знание, которое спасает жизнь
Знание, которое спасает жизнь Семь фактов о том, как глобальное потепление поджаривает США
Семь фактов о том, как глобальное потепление поджаривает США Марихуановое наступление продолжается
Марихуановое наступление продолжается
