
 От публикатора
От публикатора
Содержание
- 1 От публикатора
- 2 Стохастичность динамики среды в пространстве и пути ее преодоления
- 3 Глава 6. Функциональная организация популяции. Классификация видов с разной функциональной организацией: выделение групп и подгрупп
- 3.1 6.1. Скорость восстановления численности на депопулированных участках
- 3.2 6.2. Скорость восстановления и типы функционирования группировок.
- 3.3 6.3. Функционирование группировок и пространственно-этологическая структура: отсутствие прямой связи. Функциональные субъединицы
- 3.4 6.4. Функциональная организация и функциональная структура популяции. Изменение функциональной структуры популяции — неспецифическая популяционная адаптация
- 3.5 6.5. Виды различных функциональных групп в среде с различной динамикой
Биологи любят поговорить, что популяция это «система», в которой идёт «саморегуляция», что эволюция происходит «в популяциях», а не в выборке особей, но чаще всего это просто риторика. Для наполнения этих слов смыслом, то бишь эмпирическими данными, отвечающими на вопрос, как именно все эти общие места реализованы в природе, за счёт каких экологических и поведенческих процессов, нужно исследовать:
1) действительно ли локальные катастрофы, «точечно» ликвидирующие население вида или меняющие местообитания в неблагоприятную сторону, вызывают популяционный ответ на значительно большей площади, направленно восстанавливающий уничтоженное до прежнего уровня и/или приспосабливающий особей, «направляемых» в место катастрофы, к жизни в созданной ею экстремальной среде?
2) действительно ли население вида распределено не случайно, не ограничено лишь подходящими местообитаниями и другими барьерами, как жидкость налитая в стакан, но структурировано даже в однородной среде? Действительно ли характер этой структуризации (внешний вид группировок, изображенных «россыпью» точек в «лоскутном одеяле» местообитаний разного качества, и/или способы связанности их друг с другом) средой лишь ограничивается извне; определяется же он «конструкцией» социальной организации данного вида и популяции, их (и её) территориальными связями, всеми теми отношениями, характерный паттерн которых образует пространственно-этологическую структуру, более или менее специфичную для вида или нескольких близких видов;
3) сохраняет ли популяция устойчивость паттернов отношений, описанных в 2), как в целом, так и в отдельных группировках, вопреки «возмущениям», созданным экологической стохастичностью вне неё и демографической стохастичнстью внутри, может направленно заселять наиболее подходящие биотопы из имеющейся палитры на новых территориях и в ещё не освоенных видом ландшафтов, где среда обитания экстремальная для вида?
4) реализована ли динамическая устойчивость типа 3) благодаря общим (пространственно протяженным) регулятивным ответам на точечные (локальные) воздействия вроде описанных в 1) или нет?
Блистательные работы Н.А.Щипанова по мелким млекопитающим (Щипанов, 2000, 2003; Щипанов, Купцов, 2004) и некоторых орнитологов по синицам, дятлам, дневным хищникам (Penteriani et al, 2011; Lenda et al., 2012; Robles, Cuidad, 2017, 2020) дают положительный ответ на все 4 вопроса, наполняют фактическим материалом доселе пустую риторику про популяцию как «систему», «единицу эволюции», её «структуру» и «саморегуляцию» в ней — всё то, в чём любил упражняться Урманцев.
Всё перечисленное в кавычках осуществляется благодаря «подвижному резерву популяции» — нерезидентным особям с разной историей, направленно расселяющимся на новые территории или переселяющимся между группировками (англ. floaters). Они восстанавливают население вида на территориях, где оно исчезало после истребления человеком или природными катастрофами: легко концентрируются на вновь появившихся участках благоприятных, но временных местообитаний, интенсивно размножаются там, и также легко покидают при их исчезновении (Шилова, 1993; Щипанов, Купцов, 2004; Щипанов, 2007, 2016).
Их устойчивые потоки, «дистанционно наводимые» на подходящие, но пустующие «острова» местообитаний, или на группировки, где появились вакансии резидентов, связывают последние в системное целое, называемое популяцией, дают ей возможность восстановления после катастроф, а в спокойное время между ними — оптимального перераспределения разнокачественных особей между группировками разной устойчивости и местообитаниями разной ценности, так что их население значительно и устойчиво дивергирует между собой по признакам морфологии, жизненной стратегии, генетики благодаря данной связности, а не вопреки.
Это же верно для дифференциации населения вида в разных местностях и/или в контрастирующих ландшафтах; или для дивергенции соседних географических популяций на градиенте центр-периферия ареала, чьё значение для экологической эволюции и формообразования подробно исследовал С.С.Шварц (1980).
Самое интересное и важное в популяции не особи, разного «качества» и экологических предпочтений, а регуляторные механизмы, задействующиеся при внутренних (перенаселение или разрежение группировок) и внешних проблемах (локальные катастрофы, уничтожающие население вида в каком-то районе или благоприятные местоообитания. Это примерно соответствует разному пониманию слова рopulus как «население» (механическая сумма людей) и «народ» (единство, спаянное общей культурой, историей, традициями и сохраняющееся даже при наличии экзогамии, или смешивания с представителями других таких же единств).
Н.А.Щипанов показал, что в функциональном плане население вида в мозаике местообитаний делится на контролирующую и восстанавливающую подсистемы. Первое — резидентные особи, чья территориальность препятствует подселению новых, частот ограничивает репродуктивность уже имеющихся, второе нетерриториальные особи «подвижного резерва» популяции, активно перемещающиеся внутри неё и вовне, за границу ареала, в поисках местообитаний, сколько-нибудь подходящих, но пока что незаселенных.
«Подвижный резерв» популяции обеспечивает устойчивость её структуры вопреки гибели отдельных группировок за счёт «дистанционного наведения» особей сигнальным полем внутри видового ареала и значимыми для вида сигналами качества местообитаний вовне, там где группировки погибли из-за локальной катастрофы или вообще никогда не существовали. Первое делает кружево ареала более плотным, второе его расширяет. Соседние популяции различаются (и выделяются объективно даже в однородной среде без каких-либо преград расселению) по несинхронности этих динамических процессов. Происходящее в связи с ними в одной популяции не влияет на соседнюю и наоборот. Самостоятельность таких популяций ведёт к накоплению различий, сперва поведенческих, в виде разных традиций, поведенческих инноваций, других стереотипов связанных с местными условиями существования.
В западной литературе «популяцией» называют население данной территории, простую сумму обитающих там особей, хотя эффект Олли и прочие проявления связанности особей, управления их перемещениями и взаимодействиями со стороны целого дожны бы заставить задуматься. В отечественной, наоборот, тот же термин означает системное единство, структуру отношений, свойственных данному виду в данной части ареала, устойчиво воспроизводимых вопреки экологической и демографической стохастичности, как минимум пока численность не упадёт ниже порога минимальной жизнеспособности, и управляющих перемещениями и взаимодействиями особей на данной территории через сигнальное поле, как резидентов в группировках, так и нерезидентов в «подвижном резерве». Как именно это осуществляется у разных видов, дальше рассказывается по книге Николая Александровича «Экологические основы управления численностью мелких млекопитающих (Избранные лекции)» 2007 г.
Стохастичность динамики среды в пространстве и пути ее преодоления
Еще Д.Лэк (Lack, 1954) обращал внимание на то, что периодически обширные участки могут оказаться лишенными населения. Восстановление численности на этих площадях будет происходить за счет иммиграции особей из мест, где население сохранилось. Рассматривая численность на пространстве, занятом популяциями мелких млекопитающих, Н.П.Наумов (1945, 1948) отмечает, что существуют зоны, где гибель особей не покрывается их размножением. Пульсация пространства, заселенного видом у мелких млекопитающих, подробно рассматривается в специальной публикации В.Е.Флинта (1977), который выделяет несколько типов динамики «пространственной структуры популяций».
Обитая в динамичной среде, популяция может сохранить жизнеспособность, если она адекватно реагирует на колебания условий. Существуют модельные построения, которые показывают, что популяция, разделенная на взаимодействующие субъединицы, более устойчива к случайным воздействиям, снижающим ее численность (Свирижев, Логофет, 1978; Базыкин и др., 1980; Malchow, Feistel, 1982; Eisenberg et al., 1983). В частности, популяция одинакового объема, разделенная на две части – «ящики», связанные процессами перемещения, более устойчива к воздействию «хищника». «Хищник» может понизить численность в каждом из «ящиков» до критического уровня. В тот момент, когда он переключается на эксплуатацию другого, более выгодного «ящика», численность восстанавливается. Если популяция не разделена – «хищник» эксплуатирует ее в целом и способен понизить численность всей популяции до уровня, при котором восстановление будет невозможно (Васильченко и др., 1986).
Таким образом, для понимания устойчивости популяции важен не только анализ ее демографических возможностей, но и анализ возможных взаимодействий между внутрипопуляционными группировками (Shaffer, 1985). Локальные популяции могут обитать в разных биотопах (участках со сходными, однотипными, условиями среды). Динамика условий в разных биотопах при одинаковых изменениях некоторых факторов может оказаться разнонаправленной. Так, особи видов, требовательные к определенному уровню влажности, в засуху находят благоприятные условия в депрессиях рельефа, а при избыточном количестве осадков — на повышениях. Может оказаться, что отклонение условий в ходе случайных погодных колебаний приведет к гибели отдельных группировок. Исследуя это явление, ученые приходят к выводу о том, что гибель внутрипопуляционных единиц может возникать в различных местообитаниях и является случайной (непредсказуемой) для особей.
Если такие локальные группировки не связаны миграцией, популяция будет элиминирована через небольшой период времени (Shaffer, 1981, Гилпин, 1989, Сулей, 1989). М.Шаффер (1989) считает, что для того, чтобы сохранить жизнестойкость в реальной (гетерогенной) среде, популяции необходимо преодолеть неопределенные (случайные, непредсказуемые) влияния четырех групп факторов:
1. Неопределенность демографическая, которая порождается случайными событиями, связанными с выживанием и воспроизводством особей.
2. Неопределенность средовая возникает в результате случайных (непредсказуемых для особей) перемен погоды, обилия пищи и воздействий со стороны популяций конкурентов, хищников, паразитов и т.д.
3. Неопределенность катастрофическая, в том числе связанная с наводнениями, пожарами, засухами и прочее; такие катастрофы могут разражаться через промежутки времени неопределенной продолжительности.
4. Неопределенность генетическая порождается случайными нецеленаправленными изменениями генетического состава популяций, связанными с «эффектом основателя», дрейфом генов или близкородственным скрещиванием (с. 96).
Все эти виды неопределенных воздействий могут быть преодолены, если взаимодействие внутрипопуляционных группировок обеспечено на достаточно большой площади. Известно, в частности, что население мелких млекопитающих на прибрежных островах устойчиво поддерживается за счет периодического вселения особей (Crowell, 1973; Hanski, 1986; 1989; Peltonen, 1986; Hanski, 1991). Именно повторное заселение депопулированных участков является наиболее сложной проблемой, возникающей при ограничении численности [вредных грызунов и других хозяйственно значимых видов. Прим.публ.], поэтому, для того, чтобы добиться успеха в управлении численностью мелких млекопитающих, нам необходимо хорошо понимать закономерности пространственной динамики популяций (Krebs, 1999). Мы специально занимались этим вопросом и в следующей главе излагаем свою точку зрения.
Глава 6. Функциональная организация популяции. Классификация видов с разной функциональной организацией: выделение групп и подгрупп
6.1. Скорость восстановления численности на депопулированных участках
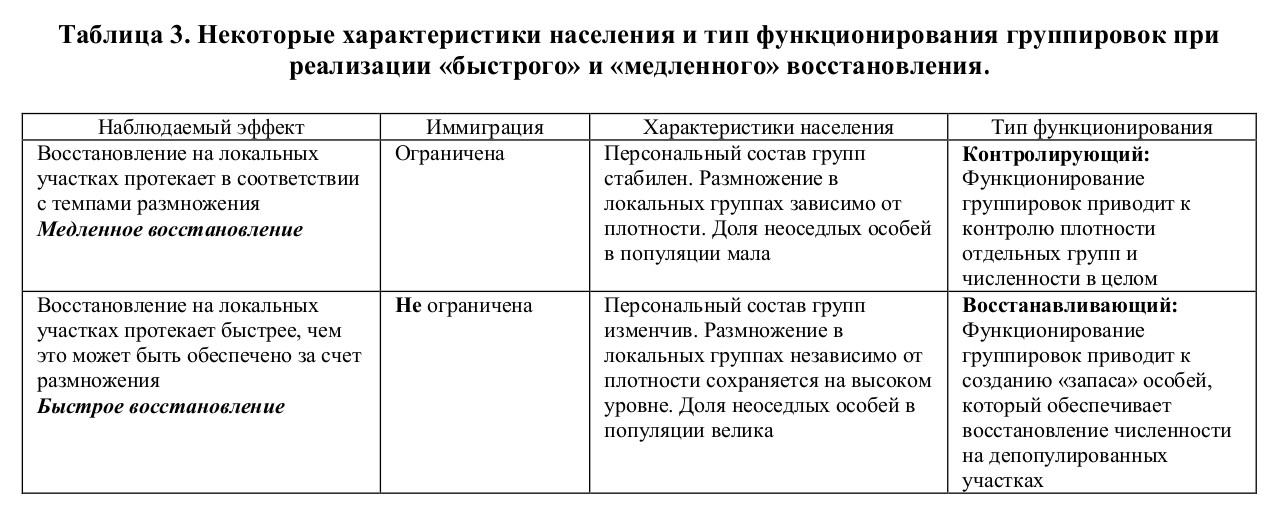
Рассматривая способность видов к восстановлению плотности на локальных участках, лишенных населения, отмечают, что восстановление исходной численности может проходить в очень сжатые сроки. Во всяком случае, быстрее, чем популяция могла бы реагировать за счет изменения темпов размножения (Анджеевский, Вроцлавек, 1962; Большаков и др., 1973). Вместе с тем, имеются виды, которые восстанавливают свою численность крайне медленно, даже при депопулировании небольших участков территории. Таковы например, полуденная песчанка и общественная полевка (Шилова, 1993; Щипанов, Касаткин, 1996; Шилова, Касаткин, 2000). Очевидно, что скорость восстановления численности у разных видов различается в деталях, однако можно указать и на принципиальное различие. Если время, потребовавшееся на восстановление локальной плотности, было меньше, чем минимальное время, проходящее от образования зиготы до созревания молодняка — минимальный репродуктивный интервал, очевидно, что восстановление численности произошло за счет иммиграции. Мы назвали такое восстановление «быстрым». Если это отношение было обратным, мы назвали такое восстановление «медленным» (Щипанов, 2000).
Очевидно, что быстрое восстановление возможно только тогда, когда в популяции имеется достаточно большой «запас» особей, которые не связаны с определенным пространством и широко перемещаются по популяционному пространству. При медленном восстановлении очевидно, что таких особей в популяции меньше, чем это необходимо для повторного заселения депопулированного участка.
6.2. Скорость восстановления и типы функционирования группировок.
Альтернативность восстанавливающего и контролирующего функционирования. Характеристики населения, связанного с обеспечением восстанавливающего и контролирующего типов функционирования
Скорость восстановления локальной плотности зависит от темпов вселения на опустевшую территорию. Поскольку «быстрое» восстановление начинается сразу вслед за гибелью населения, можно заключить, что в это время в популяции уже имеется достаточный запас особей не связанных с постоянным участком. Если количество таких зверьков в популяции недостаточно, восстановление будет «медленным». Как создается запас нерезидентных особей? Выше мы рассматривали основные принципы авторегуляции плотности населения. При насыщении локальной группы размножение должно было бы прекратится1.
Этого можно избежать, если состав группы не постоянен, а плотность поддерживается за счет баланса вселения-выселения. Соответственно, в тех случаях, когда функционирование группировок приводит к образованию популяционного «запаса» неоседлых особей, животные не имеют жесткой связи с территорией. В тех случаях, когда мы наблюдаем «медленное» восстановление, связь особей с территорией, напротив, должна быть жесткой: характеризоваться высокой степенью привязанности к освоенной территории. В этом случае торможение темпов прироста при достижении критических значений плотности является непременным условием существования, так как при избыточной плотности произойдет переэксплуатация ресурса. Таким образом, функионирование популяции, обеспечивающее быстрое или медленное восстановление противоположно по сути (альтернативно) и не может осуществляться в одном и том же месте одновременно. В общем виде основные характеристики населения, связанного с выполнением одной из функций, представлены в таблице 3.
 6.3. Функционирование группировок и пространственно-этологическая структура: отсутствие прямой связи. Функциональные субъединицы
6.3. Функционирование группировок и пространственно-этологическая структура: отсутствие прямой связи. Функциональные субъединицы
Как было показано выше (табл.3), выполнение восстанавливающей или контролирующей функции связано с такими характеристиками населения, как доля нерезидентных особей, демографический состав нерезидентного населения, и изменчивость персонального состава оседлых группировок. Это количественные характеристики. Их сходное значение может быть достигнуто за счет различного характера использования пространства оседлыми особями. Таким образом мы рассматриваем не типы пространственно-этологической структуры, а типы функционирования населения. Разные виды могут оказаться способны реализовать лишь один из альтернативных типов: контролирующий или восстанавливающий, или изменять тип функционирования в пространстве или во времени. Здесь мы рассматриваем лишь примеры изменения типов функционирования.

Изменение типа функционирования в пространстве связано с изменением значимых характеристик населения в ответ на воздействие, приводящее к снижению численности в конкретном месте. Как и в предыдущих случаях, это может быть связано с различным характером использования пространства и поведением зверьков.
Так, у домовых мышей ключевым фактором, определяющим функционирование группировок по контролирующему типу, является освоение самцами пространства. В результате образуются группировки с определенной пространственной структурой, которая поддерживается за счет охраны своей территории самцами. В насыщенной группе, образованной по такому принципу, размножение самок блокируется (Кроукрофт, 1970, Crowcroft, Rowe, 1957). Однако, если нарушить взаимосвязь особей с территорией, агрессивность зверьков падает (выше мы уже отмечали, что именно агрессивные контакты значимы для блокировки размножения) и можно ожидать восстановления размножения независимо от уровня плотности.
Это явление было экспериментально показано К.Петрусевичем (Petrusewicz, 1960). Группировка мышей с заблокированным размножением была перемещена в новую вольеру такой же площади и, несмотря на то, что плотность осталась прежней, восстановился нормальный эстральный цикл самок. Такой же эффект наблюдается и в том случае, когда группировка восстанавливается за счет повторного вселения. На освоение территории у зверьков уходит около недели. До тех пор, пока зверьки не освоили участки, они их не охраняют и в этот период на участок возможно вселение любого количества зверьков. Зверьки, не успевшие освоить участки, вытесняются, и состав группы становится нестабильным. Агрессивные контакты в такой группировке практически отсутствуют и несмотря на значительную плотность, размножение сохраняется на высоком уровне (Щипанов, Шутова, 1989). Таким образом, на месте группировок с контролирующим типом функционирования возникает группировка с восстанавливающим типом.
Общая схема изменения типа функционирования локальных группировок домовой мыши показана на рисунке 4.
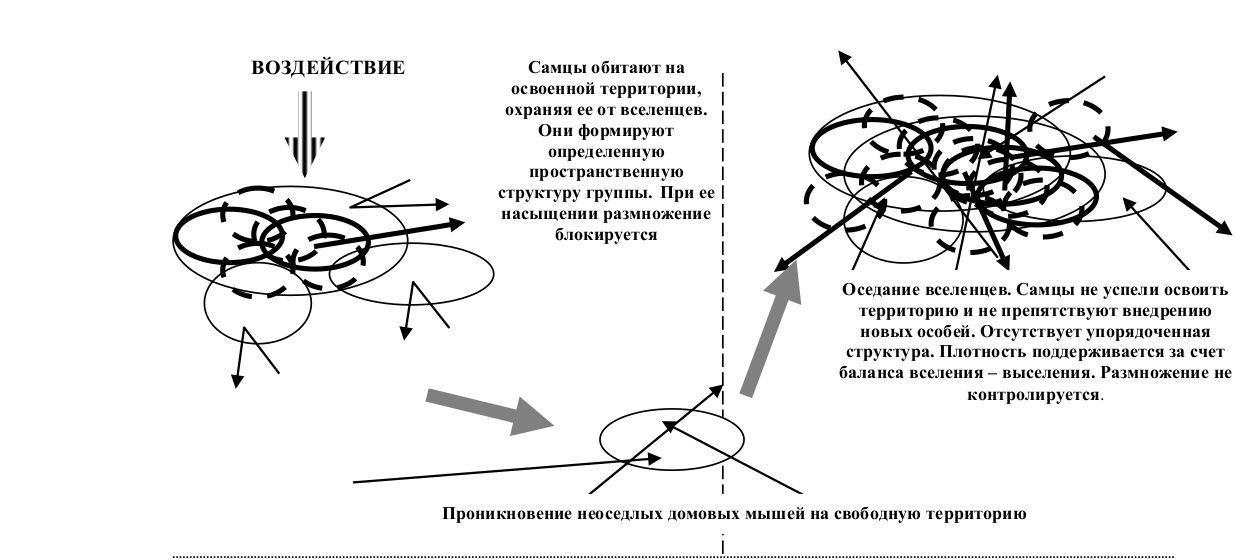
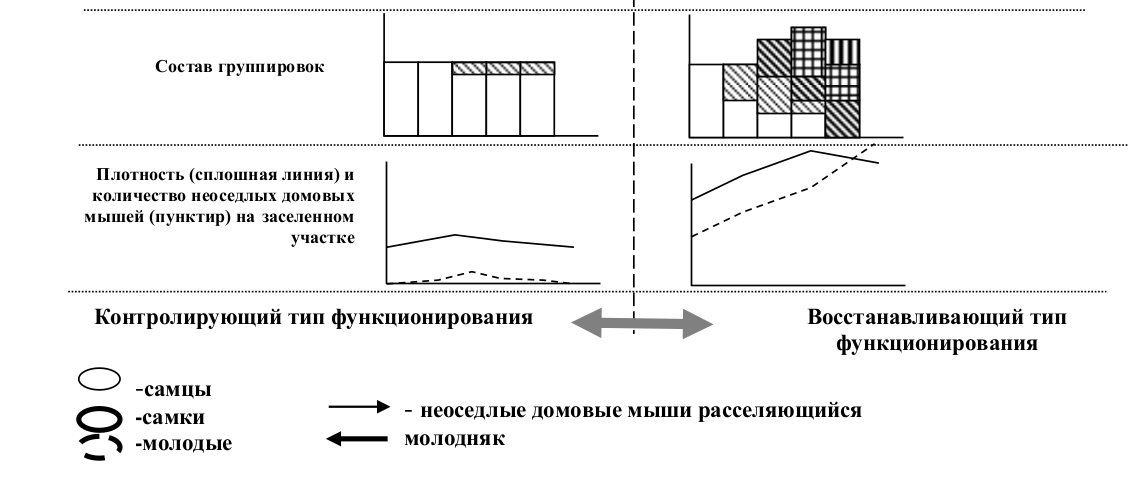 Рис.4. Схема изменения типа функционирования группировок домовой мыши в ответ на повреждающее воздействие.
Рис.4. Схема изменения типа функционирования группировок домовой мыши в ответ на повреждающее воздействие.
Изменение типа функционирования может быть основано и на других поведенческих механизмах. У лесных мышей для реализации контролирующего типа функционирования значимым является распознавание членов своей группы. Территория используется группой совместно. Чужаки изгоняются с занятой группировкой территории. При этом характерна высокая толерантность к членам своей группы и крайне высокая агрессивность к чужакам (Смирин, 1977, 1979). Для того, чтобы группировка лесных мышей сформировалась, необходим длительный период, когда отсутствует приток новых особей. Если происходит разрушение сформировавшейся группировки лесных мышей, на ее месте возникает новая, где устойчивые связи между членами группы отсутствуют. Это приводит к дестабилизации персонального состава, снятию блока размножения при высокой плотности. Отсутствие охраны территории в таких группировках способствует повышению доли неоседлых особей (Щипанов и др., 1997). В итоге группировка, возникшая на месте, где произошло воздействие, изменяет тип функционирования и начинает осуществлять восстанавливающую функцию. Обратим внимание на то, что изменение функции в последнем случае не влечет видимых изменений пространственной структуры.
Изменение типа функционирования во времени может быть связано с различным направлением отбора по признаку стресс-реактивности на разных фазах популяционного цикла. Изменение стрессреактивности особей в ходе популяционного цикла подробно рассмотрено для водяной полевки (Евсиков и др., 1995). Изменение характеристик населения, которые могут обеспечить различные типы функционирования, рассмотрены нами на примере популяции полевки-экономки на Чукотке (Щипанов, Касаткин, 1992).

При наблюдениях за зверьками в вольерах было выяснено, что группировка, образованная из неродственных особей, неустойчива. Через несколько недель выживает лишь пара. После размножения уцелевшей пары численность в группировке повышалась и в течение трех поколений устойчиво поддерживалась на уровне порядка 10 особей. То есть группировка, образовавшаяся в ходе размножения выжившей пары особей, поддерживала плотность, в 5 раз превышавшую максимально допустимую в неродственной группе. При наблюдении за популяцией полевок-экономок в природе было выяснено, что после депрессии зверьки широко перемещаются и образуют группировки, состоящие из случайного набора особей. Такие группировки характеризуются непостоянством состава, а популяция в целом — высокой долей неоседлых особей. После перезимовывания на месте этих групп остаются пары. Численность в группировках нарастает за счет оседания потомков этих зверьков. В этот период стабильность состава группировок повышается, а доля неоседлых особей падает до минимума.
На тех фазах цикла, когда популяция наращивает общую численность, она представлена группировками, функционирующими по восстанавливающему типу. Это позволяет зверькам максимально занять пригодную для жизни территорию. На этих фазах она устойчива к локальным воздействиям. На тех фазах цикла, когда наращивается плотность, группировки функционируют по контролирующему типу. Плотность и общее количество зверьков существенно возрастают, однако устойчивость локальных группировок падает. По-видимому, скорость изменений типа функционирования во времени зависит от размера взаимодействующей популяции и может оказаться различной для разных регионов.
Суммируя сказанное, мы можем заключить, что тип функционирования, как и динамика изменения функции, могут быть определены на основе анализа общих характеристик населения.
Ключевым для определения типа функционирования группировки является доля мигрантов в составе населения локальных участков. Группировки, выполняющие восстанавливающую функцию, характеризуется не только высокой долей нерезидентов, но и присутствием среди них беременных самок. Состав населения в таких группировках изменчив. Доля нерезидентов в общей структуре насления при восстанавливающем функционировании не ниже 30%, и в течение месяца обновляется не менее 20% персонального состава группы. В группировках, выполняющих контролирующую функцию, эти показатели составляют единицы процентов. Нерезидентное население представлено, в основном взрослыми самцами.
Внутрипопуляционные группировки, выполняющие восстанавливающую функцию, были названы нами «восстанавливающие субъединицы», а группировки, выполняющие контролирующую, — «контролирующие субъединицы».
6.4. Функциональная организация и функциональная структура популяции. Изменение функциональной структуры популяции — неспецифическая популяционная адаптация
Есть виды, группировки которых всегда функционируют по контролирующему типу и не способны изменить тип функционирования даже при катастрофическом снижении численности. Популяции этих видов всегда представлены только контролирующими субъединицами. Есть виды, всегда функционирующие по восстанавливающему типу. Эти виды не изменяют тип функционирования даже в ограниченном пространстве. Популяции этих видов всегда представлены только восстанавливающими субъединицами.
Некоторые виды способны изменять тип функционирования группировок в ответ на воздействие в конкретном месте. В популяциях таких видов одновременно могут быть обнаружены различные функциональные субъединицы. При этом драк между особями почти не происходит, и члены группы погибают не в результате повреждений. Можно с большой вероятностью предполагать, что зверьки погибали от стресса. Таким образом, оставшиеся полевки должны были обладать большей устойчивостью к стрессу. И, наконец, есть виды, в популяциях которых различные функциональные субъединицы могут быть обнаружены в разное время.
Представленность в популяциях вида различных функциональных субъединиц может быть рассмотрена как «функциональная организация популяции» (Щипанов, 1995, 2000). По принципу сходства функциональной организации популяции виды могут быть объединены в 3 группы и 2 подгруппы (табл. 4.).
Таблица 4. Группы видов с различной функциональной организацией популяции.
 Для видов, входящих в каждую из групп, можно ожидать определенной устойчивости и определенной динамики общих характеристик населения.
Для видов, входящих в каждую из групп, можно ожидать определенной устойчивости и определенной динамики общих характеристик населения.
Для видов 1а группы возможно изменение долевого соотношения контролирующих и восстанавливающих субъединиц — изменение функциональной структуры популяции как ответ на воздействие. Увеличение доли восстанавливающих субъединиц у 1а видов связано с увеличением пространства, подвергающегося воздействиям, вызывающим гибель населения, — повреждающим воздействиям. Повреждающие воздействия могут являться следствием различных событий, в результате которых уровень индивидуальной адаптированности особей окажется недостаточным для обеспечения их жизнестойкости: погодных аномалий, инфекции, бескормицы, вызванной деятельностью конкурентов, прессом хищников, других отклонений экосистем от «нормы», или прямых воздействий на численность, или сочетания факторов.
Независимо от специфики явлений, вызывающих гибель особей, доля восстанавливающих субъединиц в популяциях видов, входящих в подгруппу «а» 1-й группы, будет увеличиваться пропорционально территории, охваченной негативными воздействиями (рис.5). Это является адаптивным ответом популяции на любое негативное воздействие и рассматривается нами как неспецифическая популяционная адаптация. Замещение контролирующих субъединиц на восстанавливающие всегда сопровождается нарастанием доли неоседлого населения (рис.6).
Для видов, входящих в другие группы, неспецифическая популяционная адаптация за счет изменения функциональной структуры невозможна.

Рис.5. Схема изменения функциональной структуры популяции у видов 1 а группы .
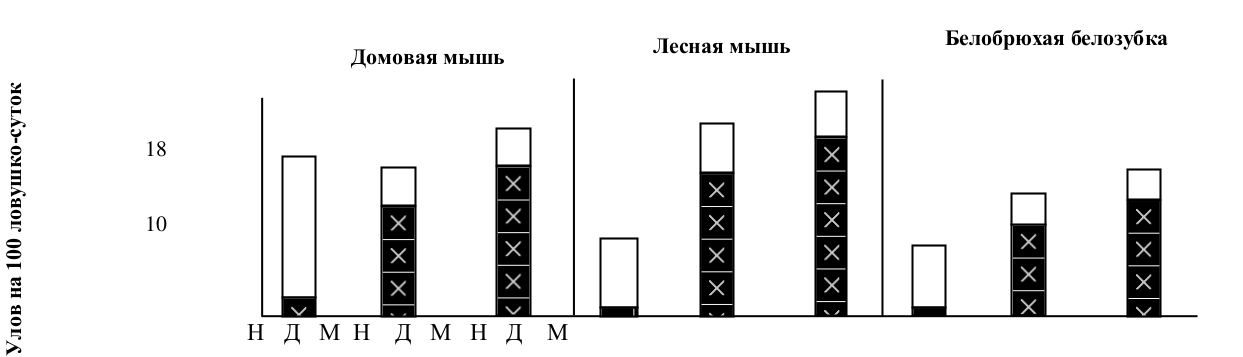
Рис.6. Численность (белый столбик) и доля неоседлых зверьков (залитая часть столбика) в популяциях видов 1а группы до воздействия (Н), через две недели (Д) и через месяц (М) после депопуляции территории.
Виды, относящиеся к 1b группе, способны изменять функциональную организацию популяции во времени. На разных фазах популяционного цикла они демонстрируют различную способность к восстановлению локальной плотности. В те годы, когда популяция представлена восстанавливающими субъединицами, эта способность высока, в те годы, когда популяция представлена контролирующими субъединицами — крайне низка. При этом доля неоседлых особей в первом случае высока, а во втором — низка (рис.7). Особенностью 1b видов является то, что на границах популяций видов этой группы может наблюдаться несинхронность популяционной динамики (Щипанов, Касаткин, 1992). Таким образом, данные, полученные для одного места, нельзя экстраполировать на регион в целом.

Виды 2 группы всегда демонстрируют «быстрое» восстановление, в популяции этих видов всегда высока доля неоседлых особей (рис. 6. 8). Они нечувствительны к локальным негативным воздействиям. Виды этой группы в норме, как правило, не достигают высокой численности, однако в нарушенных местообитаниях могут возникать концентрации зверьков со значительной плотностью. Виды 3 группы всегда демонстрируют «медленное» восстановление, а доля неоседлых особей в популяциях этих видов всегда низка (рис. 9). В норме они, как правило, многочисленны, однако неустойчивы при локальных воздействиях.
Освобожденная от видов 3 группы территория может долгое время оставаться незаселенной. Особенностью этой группы является то, что даже при низкой численности популяции плотность в уцелевших группах остается стабильно высокой.
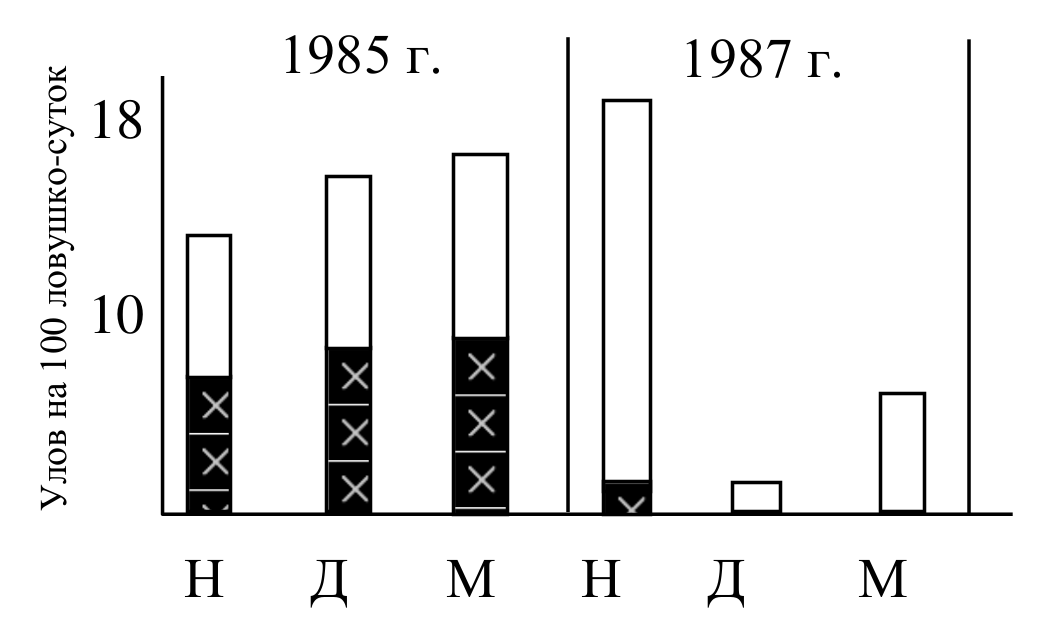 Рис. 7. Численность (белый столбик) и доля неоседлых зверьков (залитая часть столбика) в популяциях полевки-экономки – вида 1 b группы в разные годы до воздействия (Н), через две недели (Д) и через месяц (М) после депопуляции территории.
Рис. 7. Численность (белый столбик) и доля неоседлых зверьков (залитая часть столбика) в популяциях полевки-экономки – вида 1 b группы в разные годы до воздействия (Н), через две недели (Д) и через месяц (М) после депопуляции территории.
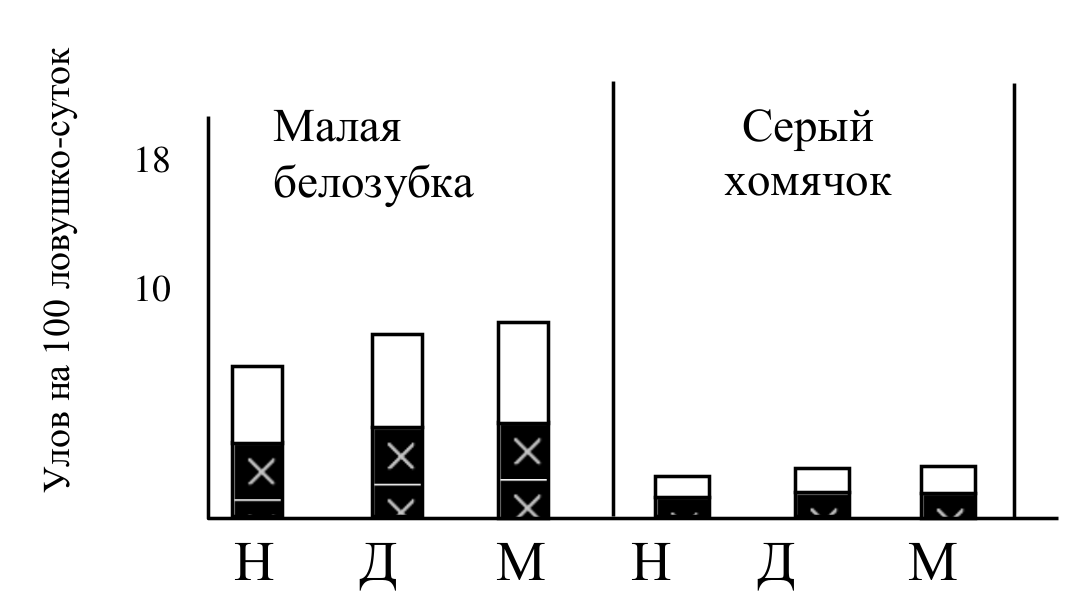 Рис. 8. Численность (белый столбик) и доля неоседлых зверьков (залитая часть столбика) в популяциях некоторых видов 2 группы до воздействия (Н), через две недели (Д) и через месяц (М) после депопуляции территории.
Рис. 8. Численность (белый столбик) и доля неоседлых зверьков (залитая часть столбика) в популяциях некоторых видов 2 группы до воздействия (Н), через две недели (Д) и через месяц (М) после депопуляции территории.
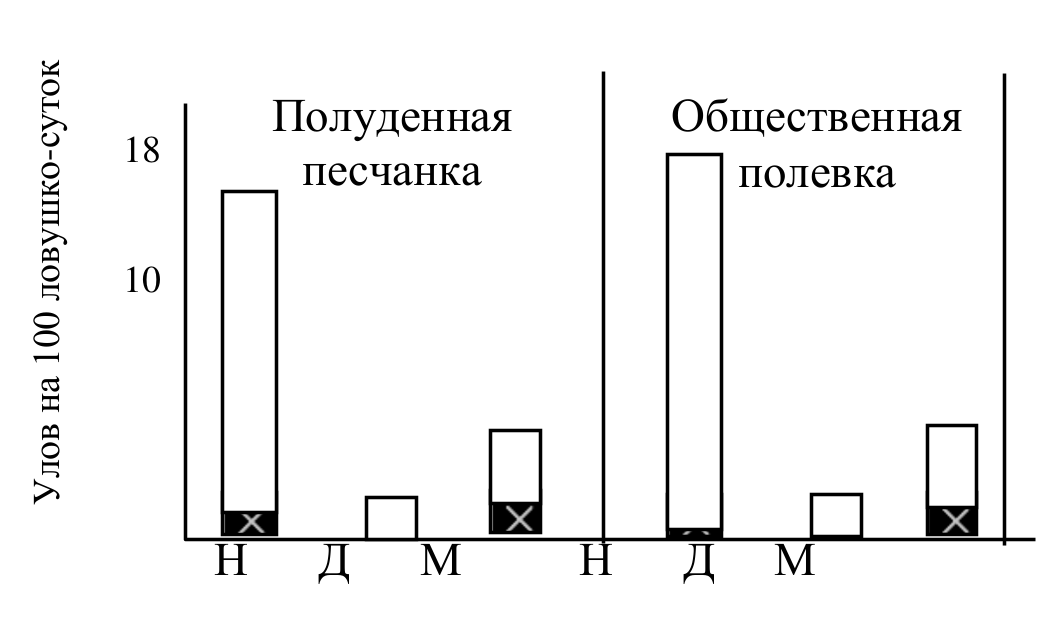 Рис. 9. Численность (белый столбик) и доля неоседлых зверьков (залитая часть столбика) в популяциях некоторых видов 3 группы до воздействия (Н), через две недели (Д) и через месяц (М) после депопуляции территории.
Рис. 9. Численность (белый столбик) и доля неоседлых зверьков (залитая часть столбика) в популяциях некоторых видов 3 группы до воздействия (Н), через две недели (Д) и через месяц (М) после депопуляции территории.
6.5. Виды различных функциональных групп в среде с различной динамикой
Все факторы, действующие на популяцию, могут быть рассмотрены с точки зрения силы их воздействия на локальных участках. Универсальная оценка силы их воздействия может быть произведена по уровню гибели особей на конкретных локальных участках.
Принципиально различным для популяции будет действие факторов, понижающих численность до уровня, когда восстановление еще возможно за счет размножения уцелевших особей — подпороговые воздействия, и до уровня, когда восстановление возможно лишь за счет повторного заселения участка — пороговые воздействия. В тех случаях, когда популяция подвержена лишь подпороговым воздействиям, для обеспечения ее жизнестойкости достаточно авторегуляторных реакций, по принципу популяционного гомеостаза в контролирующих субъединицах.

Уровень повреждающих воздействий в природе будет тем ниже, чем больше особи адаптированы к конкретной среде обитания. Однако такой уровень адаптированности индивидов одновременно приводит к тому, что их численность перестает регулироваться внешними факторами. Поэтому большое значение приобретает периодичность проявления подпороговых воздействий. В тех случаях, когда интервал между воздействиями превышает отрезок времени, за который популяция достигает критической плотности, авторегуляторные реакции во внутрипопуляционных группировках являются необходимыми.
Реализация авторегуляторных механизмов происходит в контролирующих субъединицах и, следовательно, при этих условиях все население должно быть представлено только контролирующими субъединицами. В тех случаях, когда интервал между проявлениями подпороговых воздействий меньше времени, за который популяция достигает критической плотности, плотность ограничивается без участия авторегуляторных механизмов. В условиях формирования контролирующих субъединиц не является необходимым: для обеспечения жизнестойкости достаточен любой тип функционирования.
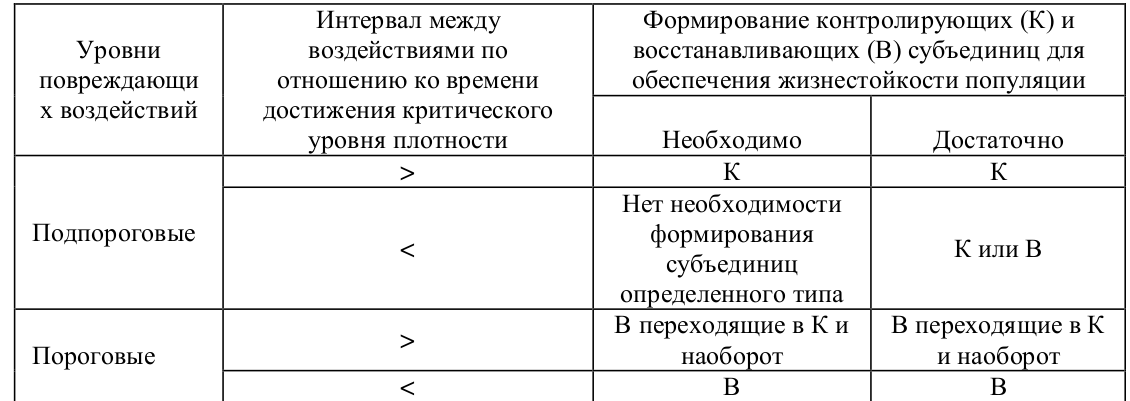
Пороговые воздействия имеет смысл рассматривать лишь тогда, когда вероятность их проявления существует в пределах всего популяционного пространства 6 , т.е. когда такие воздействия проявляются стохастически на всем популяционном пространстве. В этой ситуации необходимо наличие восстанавливающих субъединиц. Однако изменение интервала между проявлением пороговых воздействий определяет различную необходимость формирования контролирующих субъединиц.
В тех случаях, когда интервал между воздействиями больше времени достижения критического уровня плотности обеспечение жизнестойкости в благополучный период может обеспечиваться лишь контролирующими субъединицами (на тех же основаниях, что рассмотрено выше для подпороговых воздействий). Однако после воздействия жизнестойкость популяции может быть обеспечена лишь за счет восстанавливающих субъединиц. Таким образом, возникает необходимость изменения типа функционирования в различные периоды. В тех случаях, когда интервал между воздействиями меньше времени достижения критического уровня, для обеспечения жизнестойкости популяции необходимо формирование восстанавливающих субъединиц (табл. 5).
Таблица 5. Необходимость и достаточность формирования контролирующих (К) и восстанавливающих (В) субъединиц для обеспечения жизнестойкости популяции в зависимости от уровня воздействий и интервала между ними (пояснения в тексте).
Это позволяет рассмотреть возможность существования видов, входящих в разные по функциональной организации группы в среде с различной динамикой Виды обеих подгрупп 1 группы оказываются устойчивыми в наиболее широком диапазоне условий. Однако виды 1а при учащении воздействий увеличивают долю восстанавливающих субъединиц и, соответственно, долю неоседлых особей в популяции; у видов 1b увеличение подвижности происходит лишь в определенные годы.
При этом внешние воздействия на разных фазах цикла оказывают на виды 1b различное модифицирующее численность действие (табл. 6.). Виды 2-ой группы продолжают сохраняться до тех пор, пока воздействия, независимо от их величины, оказывают регулирующее действие. Виды 3-й группы сохраняют жизнестойкость лишь до тех пор, пока воздействия не достигают пороговой величины. Таким образом, для видов, входящих в определенные группы, можно ожидать и определенной устойчивости при изменении условий их существования. В противном случае ситуация не будет отличаться от рассмотренной выше, только в популяционное пространство войдут лишь те участки, где воздействия не достигают пороговой величины
Таблица 6. Допустимые области существования видов, входящих в различные по функциональной организации группы (пояснения в тексте)
 Примечание
Примечание
1Речь идет именно об авторегуляции в локальных группах. Проявление авторегуляторных механизмов для популяции в целом не препятствует созданию такого «запаса ».
Рекомендуем прочесть
 Про социальную релятивность действия окситоцина
Про социальную релятивность действия окситоцина Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов
Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов Вибрационные сигналы в коммуникации животных
Вибрационные сигналы в коммуникации животных Часть и целое
Часть и целое Стой! Не води…
Стой! Не води… Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции
Сигнальное поле как регулятор: от «памяти» и «наследственности» до эволюции Последнее слово о фетальном тестостероне
Последнее слово о фетальном тестостероне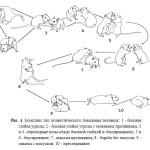 Синдром социальности: сравнительное исследование поведения песчанок
Синдром социальности: сравнительное исследование поведения песчанок
