

Резюме. В увязке друг с другом пересказаны две работы, опровергающие главные утверждения критиков сравнительной этологии, социобиологов с одной стороны, Е.Н.Панова с другой — неспецифичность мотивационной подосновы демонстраций при «семантической пустоте» их образов, предъявляемых животным партнёру (так что они не могут быть ни средством сколько-нибудь точной координации телодвижений и действий участников, ни специфическими сигналами, а тем более знаками семиотической системы данного вида, только неспецифическими раздражителями).
На деле всё прямо наоборот: молекулярными методами показана сугубая специфичность, скорость и точность мышечных движений, образующих ритуализированную демонстрацию (в данном случае барабанную дробь дятлов) в противоположность подобным им действиям, но составляющим прямые действия, скажем, кормового долбления, противопоставляемые барабанному. Неслучайно точность координации поведения участников взаимодействия — самца и самки в ухаживании, самцов, привлекающих самок и конкурирующих за них на току, на удивление высока, отражая мощь и высокую специфичность молекулярного обеспечения быстрых и точных движений мышц, описанного в этой работе.
Последнее подтверждается во второй статье, посвящённой подстановке самок-роботов самцам атласного шалашника, чтобы те за ними ухаживали. Выяснилось, что структура последовательности брачных демонстраций самца регулируется ответом самки — именно стремлением избежать разрыва в её коммуникативном внимании, чреватого тем, что она «разочаруется и уйдёт» (вызвав нешуточный стресс обоих, особенно у самца). Самец, демонстрируя, не «разогревает» самку, а «устанавливает контакты» и далее «развивает диалог», последовательно меняя демонстрации, «наращивая планку» ритуализированного ухаживания. Отсюда обязателен вывод о «семантичности» демонстраций, «работающих» предикторами поведения обоих участников взаимодействия.
Обе работы — отличный фальсификатор представлений Е.Н.Панова и других критиков сравнительной этологии (наряду с многими известными ранее), причём выполненный современными и точными методами.
***
Среди этологов есть (ложное) мнение, что демонстрации, выделяющиеся в потоке активности и составляющие этограмму, не реально существующие единицы со своей мотивационной подосновой (разные уровни агрессивной, половой и т. д. мотивации) и специфическим действием на партнёра, а преходящие моменты потока активности, которые выделимы чисто условно, в видах удобства описания с классификацией. И их появление обусловлено не спецификой ситуации вовне или мотивацией внутри животного-демонстратора (на которую они «указывают» как сигналы, соответственно референтные или мотивационные, referential signals vs motivational signals), а просто подъёмом уровня общего, неспецифического возбуждения, они как бы пена на гребне «волн» этого возбуждения, управляемых его эндогенными циклами.
У нас этот взгляд изо всех сил проповедует патриарх отечественной этологии Е.Н.Панов, пробующий подтвердить зависимость демонстраций саморекламирования черношейной каменки, потом и разных других видов от таких эндогенных циклов ещё в «Механизмах коммуникации у птиц» и ряде работ после: об организации движений хвоста ушастой круглоголовки, песни лесного конька и пр.
Однако все эти работы — подбор подтверждений тезиса, в который Е.Н. поверил заранее, а надо наоборот: сперва опровергнуть конкурирующее объяснение детерминацией спецификой ситуации/мотивационного состояния, данных о которых — хоть пруд пруди, хотя бы из уважения к критерию фальсификации Поппера. Тем более что изощрение техник этологических исследований, с ай-трекерами, самками-роботами и пр. на раз подтверждает и специфичность действия демонстраций, и следующую из этого диалоговую модель коммуникации, которую Е.Н. так долго и упорно опровергал.
Как пишет Евгений Николаевич в автобиографической книге «Зоология и моя жизнь в ней» (с.26): «уже тогда у меня зародилось сомнение в справедливости господствовавших взглядов, согласно которым коммуникация у птиц осуществляется посредством неких высоко стереотипных (ритуализованных) демонстраций, обладающих определенным сигнальном «значением». И все последующие 53 года работы и жизни в полевой зоологии он, увы, лишь подбирал подтверждения своим взглядам, как «сомнениям», так и «альтернативной теории» (с.101-108), заменяющей типологическую определённость формы и специфичность действия демонстраций1 непроверяемыми общими рассуждениями про «системность поведения» и «взаимную социальную стимуляцию». Помянув для приличия принцип фальсификации Поппера, он отнюдь им не пользуется: о массе опровержений «альтернативной теории» он просто молчит — а должен бы разобрать их и опровергат каждое по отдельности.
Недавно же Eric R. Schuppe, John O. Petersen & Matthew J. Fuxjager (2018) по барабанным дробям пушистопёрого Dendrocopos pubescens и красноголового дятлов Melanepres erythrocephalus (16 и 17 ударов в с) подробно исследовали физиологическую подоснову этих актов демонстрирования в сравнении с физиологией тех же мышц каролинского поползня Sitta carolinensis — вида, который тоже долбит (частота 5 ударов/с), но при кормлении или же расширении гнезда, а не при демонстрировании.

Пушистопёрый дятел

Красноголовый дятел

Каролинский поползень
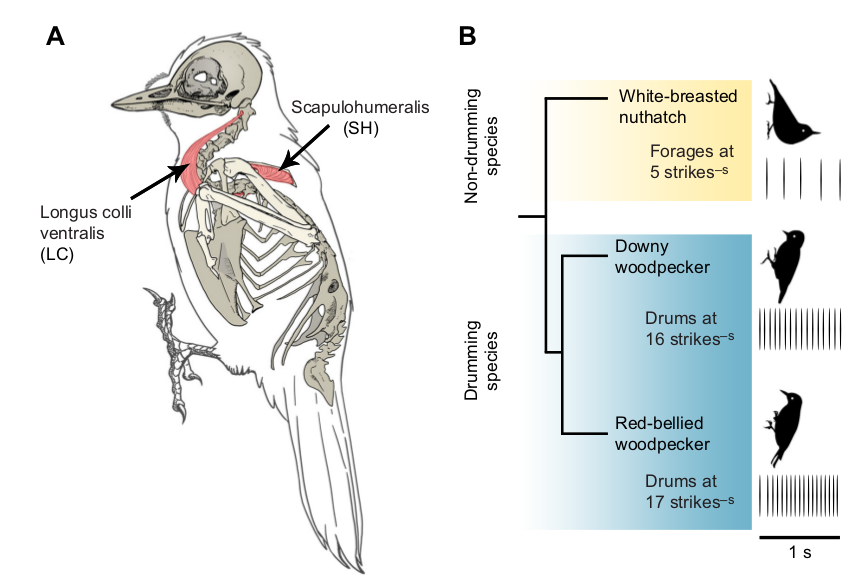 Рисунок 1. 0. Пушистопёрый и красноголовый дятлы, каролинский поползень (сверху вниз). a. Размещение изученных мышц — длиннейшей шеи и лопаточно-плечевой на скелете пушистопёрого дятла. б. Родственные взаимоотношения изученных видов, барабанящих (дятлы) и нет, но долбящих (поползень). Показаны спектрограммы барабанной дроби дятлов и кормового долбления поползня.
Рисунок 1. 0. Пушистопёрый и красноголовый дятлы, каролинский поползень (сверху вниз). a. Размещение изученных мышц — длиннейшей шеи и лопаточно-плечевой на скелете пушистопёрого дятла. б. Родственные взаимоотношения изученных видов, барабанящих (дятлы) и нет, но долбящих (поползень). Показаны спектрограммы барабанной дроби дятлов и кормового долбления поползня.
Авторы взяли две мышцы — длиннейшую шеи (musculus congus colli, Lс), непосредственно вовлечённую в «производство» дробей, и не участвующую в нём лопаточно-плечевую (m.scapulohumeralis, Sh, рис.1). В них у всех 3-х видов изучили профиль экспрессии трёх генов, белки которых регулируют транспорт ионов кальция внутри мышечного волокна, необходимый для резкого сокращения.
Это парвальбумин (PV), его белок — миоплазматический Ca2+ буфер (Celio and Heizmann, 1982), Ca2+ АТФаза в саркоплазматическом ретикулуме (SERCA1); её белок активизирует обратное всасывание иона (Feher et al., 1998; Harwood et al., 2011). Оба обеспечивают быстрые перепады концентрации кальция, позволяя мышцам ритмично сокращаться и расслабляться. Третий ген — рианодинового рецептора 1 (RYR1): кодирует белок, способствующий высвобождению Ca2+ из ретикулума. Интенсификация его экспрессии увеличивает скорость сокращения мышцы (Franzini-Armstrong, 2010).
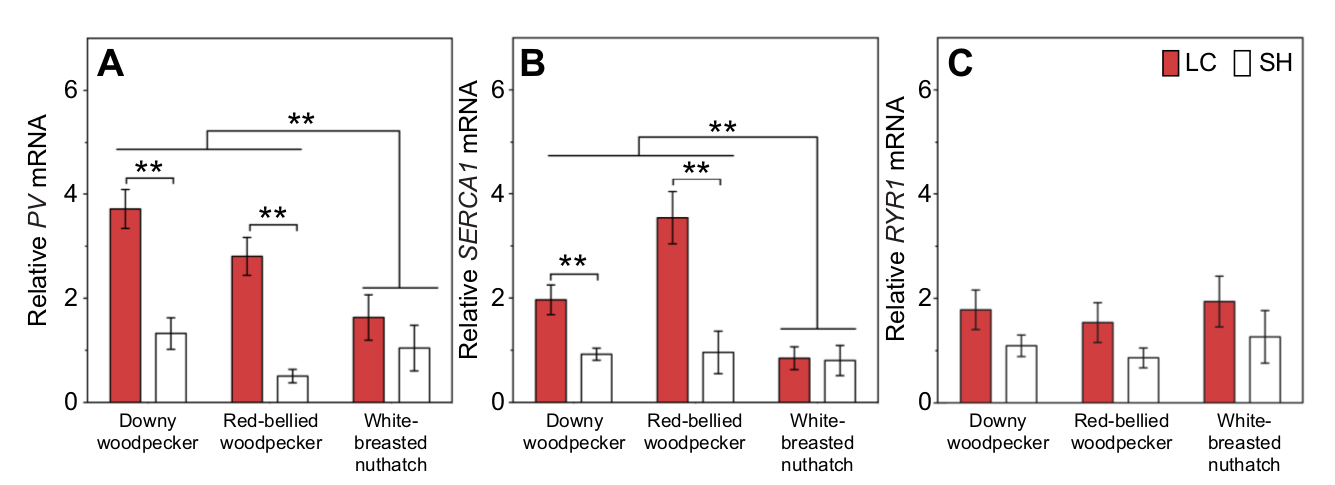 Рисунок 2. Уровень экспрессии генов парвальбумина (Pv, а); кальциевой АТФазы 1 саркоплазматического ретикулума (Serca1, b); и рианодинового рецептора 1 (Ryr1) в Lc (красные столбцы) и Sh (белые). Материал: случайно собранные в сезон размножения 8 самцов D.pubescens, 5 M.сarolinus и 7 S.carolinensis. Каждый столбец — средний уровень экспрессии в обоих мышцах данного вида, с погрешностями, отображающими ±1 s.e.m. **P < 0,01.
Рисунок 2. Уровень экспрессии генов парвальбумина (Pv, а); кальциевой АТФазы 1 саркоплазматического ретикулума (Serca1, b); и рианодинового рецептора 1 (Ryr1) в Lc (красные столбцы) и Sh (белые). Материал: случайно собранные в сезон размножения 8 самцов D.pubescens, 5 M.сarolinus и 7 S.carolinensis. Каждый столбец — средний уровень экспрессии в обоих мышцах данного вида, с погрешностями, отображающими ±1 s.e.m. **P < 0,01.
Оказывается, у обоих барабанящих видов (дятлы) экспрессия PV и Serca1, но не Ryr1, значимо выше (в 5 раз), чем у поползня, который не барабанит, именно в Lc, но не в Sh. У поползня уровни экспрессии всех трёх генов не различаются в обоих мышцах (рис.2). Буферизация же и обратный захват Ca2+ — важный медиатор быстрой мышечной сократимости, используемой именно в «складывании» и удерживании ритуализованных демонстраций. При этом повышенная экспрессия Pv и Serca1 в длинейшей мышце шеи обеспечивает барабанную дробь на молекулярном уровне, а не представляет собой его побочный продукт. Действительно, пушистопёрые дятлы барабанят примерно в 10 раз чаще красноголовых, поэтому во втором случае можно ждать, что экспрессия Pv и Serca1 вида будет в разы выше, чем у второго. Однако уровни мРНК первого гена в Lc неразличимы между таксонами, второго же выше у красноголового дятла. Следовательно, выявленный подъём экспрессии этих генов — обеспечение, а не следствие барабанной дроби, причём более эффективное у больше барабанящего вищда, ибо больший эффект достигается меньшим подъёмом экспрессии в длиннейшей мышце шеи.
Т.е. впервые так тщательно показана специфичность мотивационной подосновы телодвижений, входящих в состав демонстраций, «чертящих» их характерный облик, в противоположность того же рода движениям в составе повседневной активности. Это — отличный фальсификатор представлений Е.Н. и других критиков сравнительной этологии, наряду с многими известными ранее.
А, надо сказать, до того Eric R. Schuppe (2016) подробно исследовал охрану территории у брачных пар пушистопёрого дятла, показал, что самец и самка в противостоянии захватчикам координируют поведение друг друга, ориентируясь на вокализации, дроби и позы последнего не меньше, чем на действие партнёра (того же рода), что есть независимое подтверждение той самой диалоговой модели коммуникации и «семантичности» использованных демонстраций, визуальных и акустических, как минимум барабанных дробей, изученных авторами подробней всего, в том числе потому что их форма самая стереотипная, видоспецифическая, чётко выделимая из потока всех прочих актов животного etc.

Самец атласного шалашника в построенном шалаше; самка этого вида

Ещё раньше это выявили опыты Гейл Патричелли с соавт. с самкой-роботом атласного шалашника Ptilonorhynchus violaceus показывают, что структура последовательности брачных демонстраций самца регулируется ответом самки — именно стремлением избежать разрыва в её коммуникативном внимании, чреватого тем, что она «разочаруется и уйдёт» (вызвав нешуточный стресс обоих, особенно у самца). Иными словами, самец, демонстрируя, не «разогревает» самку, а «устанавливает контакты» и далее «развивает диалог», последовательно меняя демонстрации, «наращивая планку» ритуализированного ухаживания.

Рисунок 3. Самка-робот, подставленная самцу атласного шалашника в его шалаш. Осуществляет три типа действий, управляемых серводвигателями: приседает (опускает корпус с одновременным наклоном вперед), оглядывается при ухаживании (боковое движение головы) и расправляет крылья по достижении позиции спаривания (движение крыльев вбок).
Брачные демонстрации атласного шалашника — вычурные телодвижения, строго скоординированные между собой и закономерно развивающиеся во времени. Птица распушает оперение, затем внезапно расправляет крылья и бежит, громко жужжа. Во время каждого ухаживания самка обычно «приседает» (корпус опущен вниз и наклонен вперед в позе спаривания), при полном приседании также раскрывает крылья.
Приседания сильно варьируют у разных самок, а также в зависимости от стадии выбора самкой самца, который у шалашников сложный и многоступенчатый. Самка сперва оценивает шалаши, затем сближается с их владельцами, пробующими за ней ухаживать, затем ограничивает свой выбор одним или несколькими лучшими из их числа, и затем спаривается с каким-то одним. Приседания наблюдаются на протяжении всего этого процесса, и в целом учащаются от его начала к концу. Чем чаще приседания самки, тем большую интенсивность демонстрирование она выдерживает без риска испугом сорвать взаимодействия, когда демонстрации самца её больше стимулируют, чем пугают.
Поэтому можно считать, что приседания — сигнал отзывчивости самки: «лучший» или «хороший» самец, ориентируясь на него, выдаёт оптимальный уровень интенсивности демонстрирование, «худшие» самцы не учитывают обратной связи от самки и выдают максимум демонстрирование (или тот уровень, что присущ им per se), не считаясь с сигналами от неё. Эту гипотезу Гейл Патричелли с соавт. проверили подстановкой самок-роботов (различающихся частотой «приседаний», рис.3), в шалаши к разным самцам. Роботы облекались в шкурки самок этого вида, подставлялись в «шалаш» тестируемого самца, чтобы он за ними «ухаживал», и управлялись дистанционно. Всего протестировали 20 самцов; предложенные им «самки» различались скоростью перемещения из вертикальной позы в позу спаривания (фото) в процессе взаимодействия с самцом. Всего скоростей 4: быстрая (за 3 минуты, 1,67 смены положения в минуту), умеренная (6 и 0,83 смены в минуту), медленная (9 и 0,55 смены в минуту), и неподвижная (0 изменений в минуту). Их предъявляли случайным образом разным самцам, оценили способность «подстраиваться» к поведению искусственных самок и дальше сравнили с успехом в настоящих ухаживаниях.
Связь была сильной, линейной и положительной (рис.4), т.е. рабочая гипотеза подтверждается. В свою очередь, это говорит о роли демонстраций как сигналов-предикторов, успешно координирующих поведение участников взаимодействия, причём без этой координации успех последнего недостижим. Обратите внимание, большая интенсивность демонстраций, как это везде наблюдается у позвоночных, эффективней и рискованней одновременно. Чтобы реализовать выгоду данного поведения и не «напороться» на риск, самец должен увеличить отзывчивость на сигналы самки и/или точность координации собственного поведения с её в большей степени, чем выросла интенсивность демонстрирования.
Именно это отрицают все критики сравнительной этологии: и социобиологи, и Е.Н. Панов. Цитированные работы показывают, что они здесь опираются чисто на умозрение и игнорируют хорошо установленные факты.
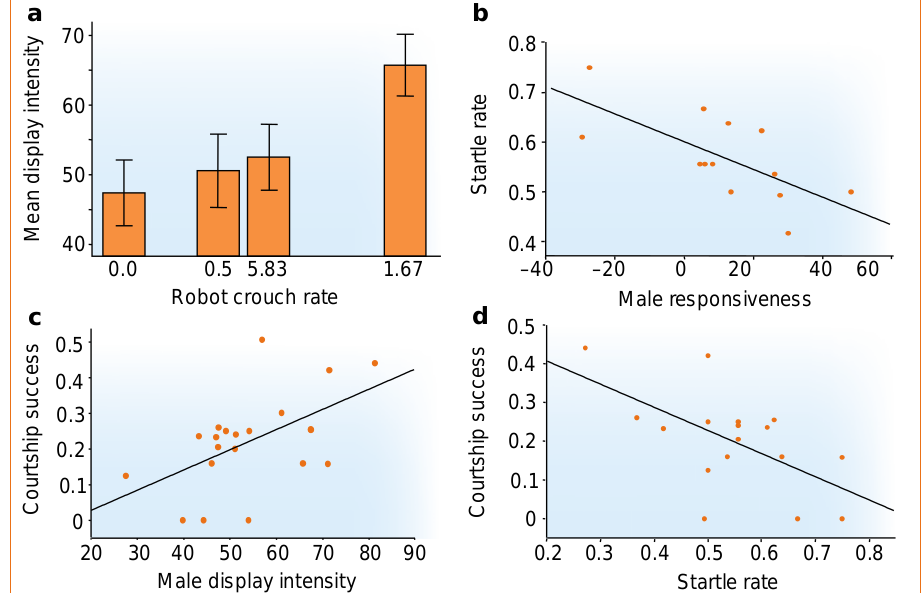
Рисунок 4. Подстраивание ухаживания самцов атласных шалашников к поведению самок-роботов. Обратите внимание на сугубую эффективность координации.
а. Средняя интенсивность демонстраций самца (5 секунд до н.э.) положительно связана с частотой приседаний «самки» (однофакторный дисперсионный анализ робота (оценка влияния фактора на отклик линейной моделью ANOVA с дискретными предикторами (контрастами), разные самцы — случайная составляющая: F1,36 =8,69, P<0,006; в оценке интенсивности брачных демонстраций учитывали аспекты телодвижений самца, создающие риск срыва ухаживания агрессией самки: распушенность оперения, длину пробежки самца и его позицию относительно самки.
б. Самцы, лучше координирующие собственное поведение с самочьим, точней реагирующие на её сигналы, меньше рисуют сорвать ухаживание, испугав её слишком интенсивными телодвижениями. Абсцисса — отзывчивость самца, ордината — риск испуга самки и срыва взаимодействия (n=13, r2=0,47, F1,11=9,93, P<0,009; отзывчивость самца определяли зависимостью интенсивности его демонстраций от частоты приседания робота, по углу наклона регрессии. Риск испуга самки рассчитан по настоящим сближениям с самками в его шалаше как частота таких случаев в ответ на интенсивные демонстрации самца в общей выборке первых ухаживаний за всеми самками, которых он туда привлёк и с которыми токовал).
в. Самцы, интенсивнее (в среднем) демонстрирующие перед роботом, успешней в ухаживаниях за настоящими самками (n=20, r2=0,29, F1,18=7,23, P<0,015; успех ухаживания самцов = доля самок, привлечённых в шалаш, с которыми самец смог спариться).
d. Самцы, реже пугающие самок слишком (для них,с учётом их собственного темпа смены форм поведения в процессе сближения с самцом) интенсивными демонстрациями, более успешны в настоящем ухаживании (n=17, r2=0,35, F1,15 =8,10, P<0,012).
Неслучайно точность координации поведения участников взаимодействия — самца и самки в ухаживании, самцов, привлекающих самок и конкурирующих за них на току, бывает высокой на удивление, отражая мощь и высокую специфичность молекулярного обеспечения быстрых и точных движений мышц, описанного выше.
Примечание
1эмпирически строго показанные: объективное знание, «добытое» сравнительной этологией, сохранилось и после того, как не выдержав волны критики 1970-1980-х, она к 1990-м годам пресеклась, будучи вытесненной из собственной проблемной области («четыре вопроса Тинбергена») конкуренткой, социобиологией. Больше того, последняя вынуждена это знание учитывать и использовать, выстраивая свои методы, хотя оно плохо «ложится» в её теории. Впрочем, в представления Е.Н. ещё хуже. Также важно, что эти данные собраны очень разными авторами, с разных теоретических позиций — и социобиологами, и «просто орнитологами«, и полевыми экологами, исследующими конкуренцию или взаимодействие «хищник-жертва».
Рекомендуем прочесть
 Хищник — жертва: психологическая «дуэль»
Хищник — жертва: психологическая «дуэль» Сколько дятлов лесу нужно?
Сколько дятлов лесу нужно? Важность повторного использования дятлами дупел
Важность повторного использования дятлами дупел Проблемы и риски выдалбливания дупел у дятлов
Проблемы и риски выдалбливания дупел у дятлов Вольфганг Шлейдт. Коммуникация животных. Поиски понимания сложности поведения
Вольфганг Шлейдт. Коммуникация животных. Поиски понимания сложности поведения Исследование маскировочности окраски дятлов
Исследование маскировочности окраски дятлов Городские ящерицы любят погорячее, и их гены объяснят, почему
Городские ящерицы любят погорячее, и их гены объяснят, почему Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов
Эпигенетическое программирование фенотипических различий в поведении и эволюция социальности у грызунов
