
 Резюме. В продолжение темы биологической подосновы борьбы с угнетением перевод статьи с результатами знаменитых опытов Дидье Дезора, про дифференциацию крыс на «добытчиков» и «эксплуататоров», многажды перевранными в Сети (даже на сайте как бы научного издательства ). Дальше два слова о современных биоинформатических подходах к поиску генов, влияющих на нашу способность доминировать и подчиняться.
Резюме. В продолжение темы биологической подосновы борьбы с угнетением перевод статьи с результатами знаменитых опытов Дидье Дезора, про дифференциацию крыс на «добытчиков» и «эксплуататоров», многажды перевранными в Сети (даже на сайте как бы научного издательства ). Дальше два слова о современных биоинформатических подходах к поиску генов, влияющих на нашу способность доминировать и подчиняться.
Авторы ставили крыс в ситуацию, когда доступ к еде был затруднён, нужно было нырнуть в водный лабиринт Томаса, плыть под водой к кормушке, далее с кормом плыть обратно. Крысы сперва приучались ходить к этой кормушке в пустом лабиринте, затем его постепенно заполняли водой, наращивая затрудненность доступа к пище. В этой проблемной ситуации крысы дифференцировались на группы с противоположными тактиками поведения, добытчиков (Д) и клептопаразитов (К); будь это люди, следовало бы писать “эксплуататоры”. Первые совершают усилие, принося пищу в общую камеру, вторые её отнимают. Самое важное из данных авторов — сугубая ситуативность этой дифференциации, она свойственна этой и только этой группе. При смене состава групп, включая создание их только из Д или К, выделяются те же самое категории особей, но положение конкретных крыс изменяется. Это куда важнее в научном и социальном плане, чем разница соотношения Д/К между двумя линиями беспородных (!) крыс, на чем концентрируются сами авторы.
Даже если они “генетические”, как они пишут, почему — непонятно, то полипотентность развития особей в сторону одной из тактик поведения и ситуативность из дифференциации в группах точно нет. Излагаются сходные данные по ранговой дифференциации в других группах позвоночных и биоинформатический анализ генов, влияющих на эту последнюю у человека. Напомним, “влиять” не значит “определять”.
Потенциальные различия в социальном поведении крыс в случае ограниченного доступа к пище
Содержание
Рэми Хелдер, Дидье Дезор и Анна-Мария Тониоло.
Анализировали социальное поведение групп из 6 беспородных крыс линий Лонг-Эванс (Лэ) и Вистар (Ви) в ситуации особенно трудного доступа к еде из-за необходимости преодолеть водный барьер и тащить еду назад в домашнюю клетку. В каждой группе лишь около 50% особей добывали пищу сами (добытчики — Д), другие же нападали на Д и отнимали её (клептопаразиты — К). Обе формы активности различаются у крыс разных линий. В линии LE, в отличие от Wi, Д приносили еду понемногу, а К этой линии были более агрессивны и чаще бывали ответно атакованы Д. Перенос корма и складывающиеся вокруг него взаимодействия, у крыс LE неизменно сложнее, чем у Wi; возможно, между линиями есть генетические различия.
Введение
Воздействие генетических факторов на разные биологические аспекты поведения показано многочисленными исследованиями на крысах, включая беспородных (для примера см. Ambrogi Lorenzini et al., 1991; Aulich and Vossen, 1978; Creel, 1980; Jori et al., 1971; Jurcovica et al., 1984; McCarty et al., 1984; Walsh, 1980). Некоторые работы сравнивали линии альбиносов Ви и пигментированных Лонг-Эванс Лэ. Скажем, Ви больше потребляют пищи, а у Лэ выше двигательная активность при беге в колесе (Ambrogi Lorenzini et al., 1987, 1988).
Однако львиная доля работ касается лишь реагирования (поведенческого или психологического) на экстремальности внешней [а не социальной] среды. Интерес же к последним растёт в связи с фармакологией. [Удовлетворяя его], авторы ранее описали социальную дифференциацию в группах крыс, чей доступ к еде ограничивал водный лабиринт Томаса (Colin and Desor, 1986; Thullier et al., 1992; Deviterne et al., 1994). Первым шагом в определении, как сильно влияет генетика на данное разделение [Д и К], было сравнение крыс ВИ и ЛЭ в этом эксперименте.
Материал и методы
В опытах использовали а) 58 беспородных самцов Ви, живших в восьми клетках по 6 в каждой, и в 2х клетках по 5. Их купили во Французской Лаборатории Селекции Иффа-Кредо; б) 53 беспородных самца Лэ до опытов жили в 8 клетках по 6 крыс и в одной по 5. Их достали во Французской Лаборатории Селекции Janvier. К моменту опытов всем зверькам было 60 дней. Температура при — 22 градуса, световой день — 12/12 ч (лампы с запасом 0700 часов). До начала эксперимента вода и стандартный корм лабораторных животных были доступны без ограничений.
Аппаратура и процесс тестирования
Экспериментальная установка показана на рис.1 (Colin, Desor, 1986) включала домашнюю клетку (50Х40Х29 см) — “базу операций” крыс, куда они возвращались с кормом и откуда стартуют, чтобы его добыть (слева). От клетки идут труба в закрытый сверху аквариум с водой (125Х20Х35 см), который надо нырнуть и плыть под водой около 1 м (в центре), чтобы попасть к кормушке (справа). Кормушка спускается прямо к поверхности воды, питание прямо в ней невозможно, нырнувшая крыса должна схватить съедобную гранулу (диаметр=10 мм, длина=25 мм, вес=2,5 г) и плыть обратно с нею в зубах.
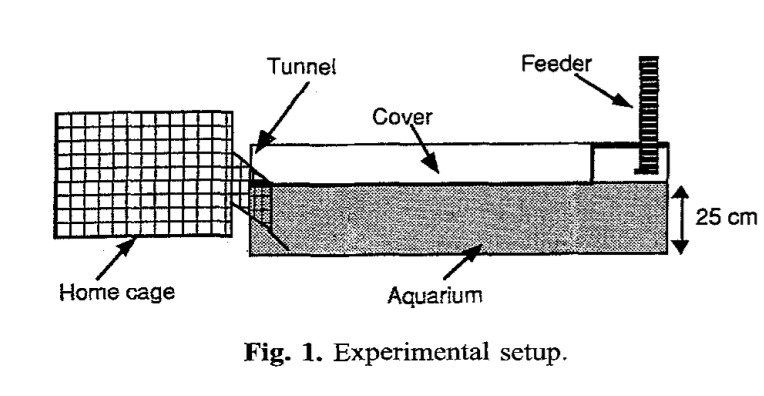 Первые 2 дня зверьки знакомились с установкой, в аквариуме не было воды, в следующие 7 дней он постепенно заполнялся так, чтобы уровень воды составил последовательно 1, 2, 5, 10, 15 и 24 см. Каждой следующей метки уровень достигал за день, и лишь последней за 2 дня. В конце концов аквариум заполнился весь (25 см). Теперь за кормом и с кормом крысы должны были плыть под водой, что не превышало 5-6 секунд, и значимо не меняло консистенцию и качество корма. Так продолжалось 21 день (температура воды 19 град.).
Первые 2 дня зверьки знакомились с установкой, в аквариуме не было воды, в следующие 7 дней он постепенно заполнялся так, чтобы уровень воды составил последовательно 1, 2, 5, 10, 15 и 24 см. Каждой следующей метки уровень достигал за день, и лишь последней за 2 дня. В конце концов аквариум заполнился весь (25 см). Теперь за кормом и с кормом крысы должны были плыть под водой, что не превышало 5-6 секунд, и значимо не меняло консистенцию и качество корма. Так продолжалось 21 день (температура воды 19 град.).
Во время опытов крыс кормили лишь 3 ч/день, без другого источника пищи. Вода была доступна по желанию. Всего использовали 10 таких установок, на которых тестировали по 5 групп Ви и Лэ ежедневно с 08.00 до 11.00, или 5 групп Ви и 4 Лэ с 13.00 до 16.00.
Наблюдения
Поведение зверьков во время опытов снимали на видео, фиксируя для каждого из них следующие переменные:
(1) число принесённых гранул;
(2) как долго они эти гранулы ели, с;
(3) сколько гранул отняли у других крыс;
(4) как долго они жрали украденное, с;
(5) сколько гранул отняли гранул у Д;
(6) сколько отняли у всех прочих крыс;
(7) сколько гранул было отнято у данной крысы К;
(8) сколько времени, потраченного на кормодобывание, каждый зверёк потерял из-за К.
Ранее авторы выяснили, что а) нормальный вес у крыс восстанавливается после первой недели с максимальным уровнем воды, б) разделение на Д и К у Ви стабилизируется к 14 дней при максимальном уровне воды, а большинство приносов гранул и, следовательно, взаимодействий между зверьками, случаются в первый час каждой сессии. Поэтому данные для настоящей статьи собирали на 21-й день, в первом часу каждой сессии, когда крысам было 90-95 дней. У всех них поведение фиксировал один и тот же обученный наблюдатель.
Анализ данных
Большинство поведенческих переменных имеют распределение, отличное от нормального. Поэтому для оценки усредненных значений переменных и их изменчивости использовали медиану, нижний и верхний квартили. Поэтому же значения переменных для Ви и Лэ сравнивали с использованием непараметрической статистики, критериев хи-квадрат и Манна-Уитни.
Результаты
Для взятия корма крыса должна проплыть под водой, а чтоб его съесть — снова плыть, возвращаясь. Среди крыс ВИ так ведут себя только Д, остальные (К) у них корм отнимают. В данном исследовании разделение на Д и К обнаружено также у крыс Лэ. Из 58 крыс Ви 28 (48.3%) были Д, из 53 Лэ — 27 (50.9%), различия не значимы (X 2 = 0.01; NS).
Поведенческий профиль Ви
Представлен на рис. 2-3. Так, обобщëнный Д за первый ч опыта 8,5 раз нырял за гранулой и обратно (рис. 2а-f). Сам он, в общем, не отнимал еду у других, питаясь только плодами своих трудов, на что ушло в общем 1538 с. Только половину корма они умели сохранить для себя: из 8,5 гранул 4 были отняты К, те за их счёт кормились 1245 с.
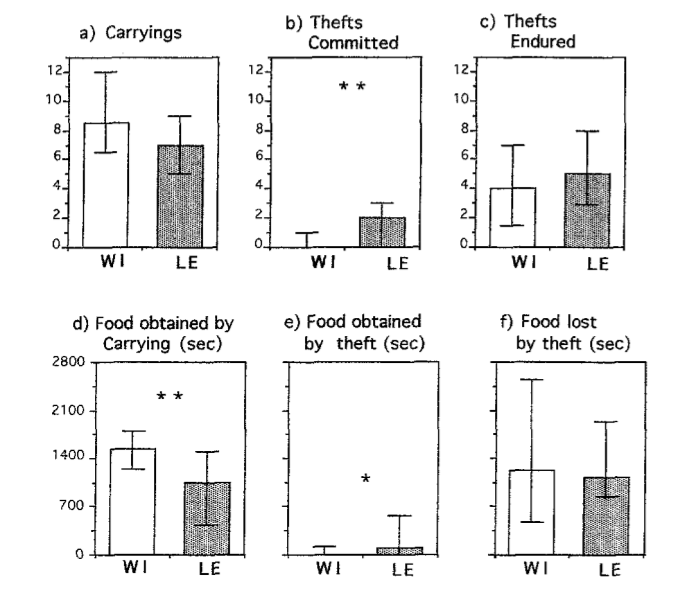
Рис. 2. Различия Ви и Лэ по фиксируемым поведенческих переменных: а) число принесённых гранул, b)-c) число совершенных и перенесенных краж, d)-e) время кормления происходившего после приноса корма самому и отъезда чужого, f) потерянное время кормления из-за К (f). Приведены медиана, верхний и квартили. Значимость различий — критерий Манна-Уитни, (**) p < 0,005; (*) p <0,05.
Прочие особи (К) вели себя совсем иначе (рис. 3a-d): отнимали еду у Д (медиана успешных отъëмов = 5), но сами такому не подвергались (медиана отнятых у них капсул = 0). ¾ из них никогда не отнимали еду у других К, только у Д. Тё же наоборот, если и отнимали еду (очень немного), то лишь у других Д.
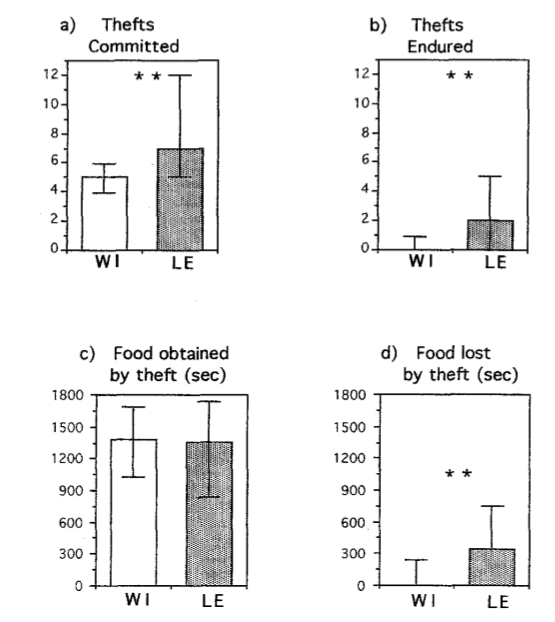
Рис. 3. Сравнение (по Манну-Уитни) количества совершенных и перенесенных отъемов корма у лиц, не являющихся Д, из пород Ви и Лэ (а, b), и времени кормления, полученного в результате кражи (c) или потерянного в результате кражи (d). Данные представлены в медианах и квартилях. (**) р <0,001.
Рис.4 показывает % отъема пищи внутри групп и между группами (стрелки) к общему числу отъëмов за время наблюдений (а — Ви, b — Лэ). Основная масса взаимодействий (87,8% у Ви, 53,4% у Лэ) происходит между Д и К, отнимающими у них пищу. Оставшиеся (12,2%) делятся на 3 категории: между К (5,8%), между Д (4,5%) и самое редкое — Д отнимают еду у К (1,9%).
Поведенческие характеристики Лэ
Показаны на рис. 2а-f. Здесь типтчный Д нырял и возвращался с кормом 7 раз, питался он в общей сложности 1045 с, ещё 108 с питания у них добавлялось за счёт 2х успешных отъëмов пищи. В свою очередь, у него еду отнимали 5 раз, из-за чего он терял 1118 с. возможного времени кормления.
Прочие крысы Лэ (т.е. К) всю пищу добыли в ходе взаимодействий с другими зверьками (рис. 3a-d). Они 7 раз отнимали пищу, получая потенциально 1356 с кормления, но 2 гранулы (341 с времени) теряли за счёт отъемов, совершавшихся как Д, так и К.
Типичный К линии Лэ 4,5 раза отнял пищу у Д, 1,5 — у другого К (“вор у вора дубинку украл”, 18,7% случаев). Даже Д отнимали пищу у других Д (10,6%), и даже у К (17,3%), см. рис.4b.
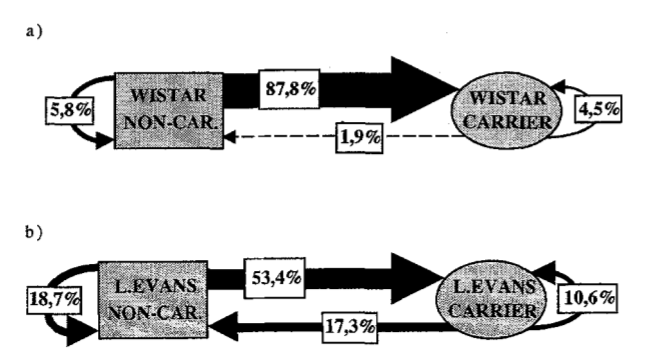
Рис. 4. % отъемов корма внутри и между разными категориями крыс (Д vs прочие) у линий Ви (a) и Лэ (б). Стрелка идёт от К к его жертве.
Таблица I сравнивает по критерию Манна-Уитни частоты отъëмов пищи у Д и К из линий Ви vs Лэ. Значимыми оказываются различия между линиями в 3х комбинациях из 4.

Сравнение поведения Ви и Лэ
Рис. 2a-f показывает, что интенсивность кормления у обоих линий значимо не отличалась. Обе испытывали одинаковое “давление” клептопаразитизма (U = 294, NS) и добывали равное количество корма (U : 351, NS). Что отличалось, так это активность Лэ: они, включая Д, совершали больше отъемов пищи, чем Ви (U = 313,5, p < 0,05), чем компенсировали меньшую эффективность доставки гранул (U — 195, p < 0,005). Они также чаще взаимодействовали друг с другом (рис. 3a-d), совершив в 2 раза больше отъëмов пищи (U = 221,5, p < 0,001). Однако это не пошло им на пользу, время кормления не выросло (U = 370,5, NS). Данные К значимо чаще делались жертвами других К, чем в случае Ви (U = 187, p < 0,001). Эти потери пищи они компенсировали ростом своих отъëмов (U = 208, p < 0,001) и т. д.
Анализ сети взаимодействий Лэ и Ви в табл. 1 показывает, что рост числа отъëмов корма у Д Лэ в равной мере направлен на К и других Д. Хотя в среднем К также часто отнимали пищу у Д, рост частоты взаимодействий у Лэ вëл к тому, что каждая гранула “переходила из рук в руки” по нескольку раз, в этом участвовали 3 или 4 особи. Напротив, у Ви корм или оставался у Д, или один раз отнимался К.
Обсуждение
Пища в экспериментах приносилась лишь Д, доля которых сравнима в обеих линиях (49% у Ви и 51% у Лэ). Однако перераспределялась она сильно по-разному. Подвергшись отъëму пищи, Д линии Ви практически не показывали ответной агрессии, но интенсифицировали доставку пищи. Соответственно, К до блеска отточили отъëм, Д им не пробовали сопротивляться. У Лэ было иначе, потерявшие корм крысы сами в ответ отнимали его у любых обладателей корма, Д или К. Однако у Д агрессия подавлялась не полностью, лишь временно. При формировании групп крыс заново, только из Д, они дифференцировались на Д и К без сохранения прежних ролей (Colin, 1989). Напротив, у Лэ агрессия Д не подавлена полностью нападениями К, в ответ они сами отнимают пищу у любых других крыс. К здесь менее эффективны в охране отнятой пищи, что увеличило частоту конфликтов (341 против 156 у Ви), при учащение перехода гранул “из рук в руки”.
Таким образом, данное исследование продемонстрировало потенциальное генетическое влияние на социальное поведение крыс в ситуации ограниченного доступа к пище. Следующим шагом в этом направлении, по-видимому, будет проверка на наличие различий и/или сходства внутри штамма, по методу Файлаа и Веллуччи (1979), с использованием Ви и/или Лэ-крыс, которые были предоставлены разными поставщиками. Следующим шагом был бы контроль возможных внутригрупповых различий у крыс Лэ путем тестирования каждой группы в течение нескольких дней.
У людей отбор равно поддерживает гены, влияющие на тенденцию доминировать и подчиняться
Недавно новосибирские биоинформатики (Ирина Чадаева с соавт., 2019) подошли к той же проблеме с другой стороны, и для самого интересного вида — нашего собственного. Они предложили эвристическое правило поиска генов, влияющих на социальную дифференциацию у гомосапов, т.е. разделение на доминантов и подчинённых в разных системах иерархии, ему присущих, в каждой идут конкурентные столкновения, делящие участников на победителей и побеждённых со значительным последействием, накопление их результатов и делает одних доминирующими, других подчиняющимися, но лишь в данной группе и на данном поприще.
Это верно и для животных, почему индикаторы агрессивного поведения и социального доминирования изучали на мышах, отселектированных согласно методе Д.К.Беляева на миролюбие, или наоборот, агрессивность, с выявлением генов во всём этом участвующих. Мышам создали историю повторяющихся побед и поражений в стычках при ссаживаниях. Для этого к более сильной особи подсаживали заведомо более
слабую — или наоборот, давали им драться по 5 минут, после чего повторяли; и так до тех пор, пока не сформируется устойчивый, но не наследуемый комплекс поведения победителя (наглый, задиристый, легковозбудимый, почему охотно лезущий в драку) и проигравшего (неофобный, всего боящийся, пассивно избегающий драк).
Это изменения модификационные, но крайне устойчивые, и можно выявить гены, чьи
изменения экспрессии участвуют в создании этой устойчивости, закреплении каждого из психотипов. Сходная дифференциация психотипов при победах и поражениях в стычках отмечена и у лабораторных крыс. Собственно, это норма для агонистического поведения позвоночных: выигранные и проигранные конфликты имеют своё последействие, специфически увеличивающие вероятность наступление того же исхода в следующем взаимодействии, пусть даже с другой особью, и требующие использования более эффективных демонстраций для его изменения в следующий раз.
Возвращаясь к сказанному: правило поиска генов-участников таково: ЕСЛИ {избыток белка, связанного с нервной системой — экспериментально известный физиологический маркер низкой болевой чувствительности, быстрого восстановления после конфликтов, или аналогичных им форм агонического / интолерантного поведения, вроде спортивных состязаний} И ЕСЛИ {данный одиночный нуклеотидный полиморфизм (SNP) вызывает сверхэкспрессию гена, кодирующего этот белок} ТО {этот SNP может быть маркером SNP тенденции к доминированию} ПОКА {недоэкспрессия того же самого гена соответствует субординации} И наоборот.
На основе данного правила они изучили 231 ген Homo sapiens из нейропептидергической, ненейропептидергической и нейротрофинергической систем. Там они кодируют нейротрофические и ростовые факторы, интерлейкины, нейротрансмиттеры, рецепторы, транспортеры и ферменты: соответствующие белки известны как ключевые факторы социального поведения человека (в смысле участники «исполнения» разных форм поведения, выбранных и осуществляемых нами, частью инициативно, частью в ответ на внешние воздействия, которые почти исключительно символы культуры, включая язык, прямые действия, вроде болевых, температурных и сексуальных, их лишь сопровождают). Точнее, анализировались не сами гены, а системы управления ими — промоторы, часто находящиеся далеко от них по «тексту» ДНК, и использующие ТАТА-связывающие белки для регуляции, усиливающей или ослабляющий экспрессию соответствующих генов.
Авторы проанализировали все 5052 SNP в промоторной области размером 70 п.н. выше позиции, где начинается транскрипт «текста» каждого гена, кодирующий его специфический белок. Анализируемые области ДНК были извлечены из баз данных Ensembl и dbSNP с помощью ранее созданного ими общедоступного веб-сервиса
SNP_TATA_Comparator.
Это определение промоторной области включает все ТАТА-связывающие сайты.
Всего найдено 556 и 552 SNP-маркёров, кандидатов на участие в реализации доминирования и подчинения соответственно. На этом основании авторы определили, что данные 231 ген подвергались естественному отбору против недостаточной экспрессии (значимо p < 0,0005).
Т.е. доминирование и подчинение равно оказываются частью нормы реакции, отражающей пластичность социальной системы человека, и отбор поддерживает тенденции к тому и другому, исходя из обстоятельств столкновений, которые оказываются важнее всего: а они социальны, «биология» же (изменения экспрессии генов, за ними гормонов, настроения, состояния и проч.) образует исполнительную часть реализации социальных влияний в поведении. К слову, такой двунаправленный отбор по генам, вовлечённым в осуществление социальной иерархии, даёт независимое подтверждение известной гипотезы самодомашнивания человека.
Рекомендуем прочесть
 Стресс в раннем онтогенезе как адаптивное явление
Стресс в раннем онтогенезе как адаптивное явление Как иерархия в группе влияет на её эффективность?
Как иерархия в группе влияет на её эффективность? Про социальную релятивность действия окситоцина
Про социальную релятивность действия окситоцина Крысы и право собственности
Крысы и право собственности Лабораторные крысы
Лабораторные крысы Борьба с угнетением: биологический бэкграунд
Борьба с угнетением: биологический бэкграунд Социальный ранг и психика. Синантропные грызуны в «жилой комнате»
Социальный ранг и психика. Синантропные грызуны в «жилой комнате» «Разумная предусмотрительность» и другие особенности поведения врановых
«Разумная предусмотрительность» и другие особенности поведения врановых
