

Самец и самка кассиева клеста у шишки сосны скрученной
Cody K. Porter and Craig W. Benkman
Культурная эволюция [точней, социальная трансляция выучиваемых признаков, одновременно воспроизводящая сходства внутри популяции и отличия от соседних, буде они появятся. Культуры в человеческом смысле у животных нет, даже у антропоидов или попугаев. Здесь и далее прим.перев.] может ускорить дифференциацию популяций и [в конечном счёте вести к] видообразованию. Правда, большинство систематиков под последним по умолчанию имеет в виду аллопатическое видообразование, когда механизмы социальной трансляции [она же сигнальная наследственность — всё что воспроизводится в нисходящем колене через обучение и близкие к нему механизмы, вроде импринтинга] закрепляют и воспроизводят случайные различия, накапливающиеся в изоляте за время его отдельного существования [культурный дрейф].
Гораздо меньше внимания уделяют роли этого механизма в иных формах видообразования, когда изоляция между обособляющимися формами отсутствует, между ними всё время разделения, достаточно долгое, сохраняется поток. Одна из клад (филогенетических ветвей), в которой реализована последняя ситуация — группа клестов-еловиков Loxia curvirostra, распадающаяся на множество форм, часть которых может быть названа экотипами, но все они представляют собой коммуникативные сообщества, отличающиеся друг от друга вокализацией, в том числе и в условиях симпатрии.
Cтатья Крэйга В. Бенкмана, много лет плодотворно изучающего формообразование у еловиков, «Смещение признаков [вокализаций клестов] в ходе социального обучения и его следствия для экологического [то бишь симпатрического] видообразования» посвящена клестам Южных Холмов (штат Айдахо), исходно выделенным как экотип [самый крупно- и толстоклювый, специализированный к добыче семян из шишек сосны скрученной широкохвойной], и только недавно получившим таксономический статус, даже видовой (Loxia sinesciuris). Как и у прочих клестов, позывы, используемые в социальных контактах, молодые выучивают от родителей.
Прошлые исследования показали неслучайное изменение птицами своих позывов на протяжении жизни: вектор динамики направлен на рост их отличий от экотипа, близкородственного sinesciurus, и симпатричного с ним [он специализирован к добыче семян из шишек сосны жёлтой подвида Скалистых гор, Pinus ponderosa scopulorum] (рис.1).
Это фактически настоящее смещение признаков между обоими формами (character displacement): о нём много говорят систематики применительно к случаям аллопатрического видообразования, однако оно фатально отсутствует там, где его ждут, при вторичном контакте частично разошедшихся и достаточно долго изолированных пространственно форм. А там, где не ждут, при симпатрическом обособлении форм в контрастирующих обстановках за счёт разделения их как коммуникативных сообществ, такое встречается, чему и посвящена данная работа.
Подобное разделение увеличивает приспособленность: расхождение вокализаций минимизирует риск присоединения к стае «чужой» формы (неадаптивного, ибо снижает эффективность кормления). В использовании запасов корма (шишек хвойных) кресты ориентируются на социальную информацию [как ведут себя другие особи, останавливаются кормиться на данном деревня/группе деревьев или, минуя их, летят дальше, англ. Social information]. Поэтому смешанные стаи из птиц разных форм кормятся менее успешно.
Авторы подтверждают устойчивое распознавание «своих» и «чужих» между обоими симпатрическими формами еловиков, и смещение признаков, делающее его более эффективным. Ими фиксируется значительная дивергенция в структуре вокализаций клестов Южных Холмов, случившаяся всего за два десятилетия; также проведены полевые эксперименты, показывающие устойчивое распознавание различий в попытках и реагирование на них объединением «со своими» клестами при избегании «чужих».
1. Введение
Содержание
Когда-то культуру считали уникальной особенностью человека, но в последние десятилетия [появилось] все больше данных о культуре [1] и культурной эволюции [2] в животному мире. [На деле это не культура, а просто сигнальная наследственность, она же социальная трансляция: воспроизводство приобретённых признаков через обучение действительно важно в выработке адаптаций. Используя подходящую здесь терминологию кладизма, можно сказать, что пишущие о «культуре животных» выделяют культуру по плезиоморфным признакам (передача возникшей новизны обучением), а надо по синапоморфным (творчество, т.е. направленное генерирование такой новизны, стимулируемое социумом, благодаря чему не приходится ждать её появлений как редких событий)] [3].]
Исследования последних десятилетий показали, что культурная передача влияет на эволюцию кормодобывания [4], использование среды обитания [5] и миграцию [6]. Некоторые исследования предполагают, что культурная эволюция этих и других форм поведения может взаимодействовать с эволюцией на уровне генов и даже стимулировать ее [7,8]. Сильнее всего первая может влиять на вторую при отщеплении «дочерней» формы от «материнской» (либо ином их разделении в процессе видообразования), когда поток генов между расходящимися линиями сокращается и в конечном итоге устраняется [9]
[Из-за того, что социальная трансляция обеспечивает распознавание «своих» и «чужих», с предпочтением первых и отвержением вторых. Далее этот механизм аффилиации-дискриминации обозначается МАД]. Быстрая эволюция признаков, передаваемых сигнальной наследственностью, влияет на репродуктивную изоляцию (формы брачных сигналов и предпочтения определённых фенотипов как партнёров). Поэтому вероятно, что обучение может способствовать быстрому расхождению сигналов и предпочтений между популяциями [1,10–13]. Действительно, сейчас есть достаточно эмпирических данных, как культурная эволюция способствует аллопатрическому видообразованию ([11,14–16]; но см. [17]), особенно при культурном дрейфе — случайном расхождении между популяциями по выучиваемым признакам [15,18,19] — а также теоретических моделей, показывающих, как это происходит.
Однако намного меньше свидетельств роли культурной эволюции в случаях симпатического видообразования, когда изоляции нет, а поток генов сохраняется весь период разделения форм (обычно это экологическое видообразование, когда разные генеалогические линии «разрывает» дивергентный отбор [20]). Фенотипы таких линий дивергируют друг от друга, так что отбор может способствовать выработке отдельных сигналов, связанных с каждым из них (т.е. маркеров, отделяющих один фенотип или группу от другого/ой). Её приспособительные значение — в снижении вероятности смешения (объединения в стаю, образования пар и т. д.) между линиями ввиду его неадаптивности. Если такая дивергенция признаков «для распознавания» идёт быстро, это и есть упомянутое «смещение», character displacement [13,21–23]. Если такие маркеры групповой принадлежности влияют на выбор партнера (а тем более если он основан на использовании этих сигналов), расходящиеся линии делаются отдельными коммуникативными сообществами и становятся изолированней репродуктивно, с усилением дивергенции по подобным маркерами, дополнительно «разрывая» группы, уже разделяемые дизруптивным отбором [23,24].
Когда дивергирующие сигналы и на их основе — МАД передаются негенетически, выучиванием от родителей, потоку генов и рекомбинациями не разрушить установившейся корреляции между определёнными формами маркерных признаков (контрастирующими в разделяющихся сообществах) и предпочтением их «своими»/отторжением » чужими» особями. Это устраняет основную помеху симпатричнскому видообразованию — поток генов, потенциально нивелирующий идущую дивергенцию [24]. Однако обучение может работать и против него, ведя к конвергенции сигналов через выучивание «чужих» и успешное пользование ими в коммуникации; здесь поток генов, наоборот, увеличивается [24,25]. Какой вариант реализуется в каждом конкретном случае, из общих соображений неясно [26].
В настоящей работе отслежена долговременная эволюция выучиваемых позывов еловиков; также экспериментально проверено, обеспечит ли дивергенция таких признаков, как структура позывов, репродуктивную изоляцию между разными формами клестов-еловиков, питающихся семенами хвойных.
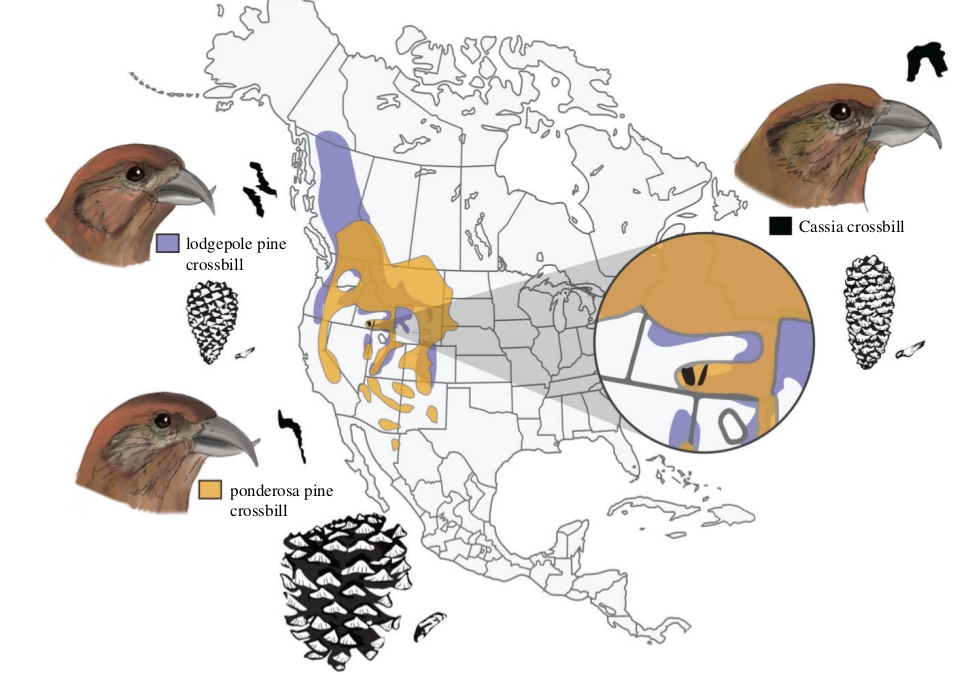 Рисунок 1. Ареалы разных экотипов («племен») еловика. Фиолетовое — клесты, специализированные к сосне скрученной, оранжевое — к сосне жёлтой или орегонской. Оба они симпатричны с узкоареальным клестом Южных холмов, или кассиевым (на врезке чëрным). Также изображены головы самцов всех 3х экотипов, предпочтитаемые ими шишки и семена хвойных деревьев, и спектрограммы контактных позывов всех трёх форм.
Рисунок 1. Ареалы разных экотипов («племен») еловика. Фиолетовое — клесты, специализированные к сосне скрученной, оранжевое — к сосне жёлтой или орегонской. Оба они симпатричны с узкоареальным клестом Южных холмов, или кассиевым (на врезке чëрным). Также изображены головы самцов всех 3х экотипов, предпочтитаемые ими шишки и семена хвойных деревьев, и спектрограммы контактных позывов всех трёх форм.
Шишки сосны скрученной также годятся для клеста Южных Холмов, но они лучше защищены от вскрывания, чешуи крупнее и толще. Соответственно клюв этих клестов выше и толще, вследствие коэволюции с шишками местной сосны.
В Северной Америке комплекс клестов-еловиков (Loxia curvirostra) состоит из девяти экотипов, вполне симпатичных, обособившихся благодаря дизруптивному отбору по форме и размеру клюва в связи со специализацией к питанию семенами разных видов хвойных [которые добывают из шишек разного размера и толщины чешуй][27]. У разных экотипов также различны акустические структуры контактных позывов, обмен которыми сплачивает стаи (в них птицы проводят большую часть жизни, рис. 2а; [29]) и стимулирует общение её членов (рис.2с; [28]). Эти различия контактных криков могут быть маркером принадлежности к одной из 9 форм, облегчающим приспособительное объединение в стаи птиц только одной формы, с одинаковой формой клюва, т. е. ассортативности [30,31]). Для определения качества кормового дерева (стоит его покинуть или можно кормиться дальше) клесты используют как свою собственную скорость потребления семян [из тамошних шишек], так и аналогичную скорость поступления корма у сочленов по стае, т. е. социальную информацию [31].
Однако ориентация на неё повышает эффективность кормления лишь если все клесты в ней примерно одинаково обрабатывают шишки и достают семена, т. е., относясь к одному экотипу, имеют в среднем одну форму клюва [27]). И наоборот: эффективность кормления снижена, когда стая смешанная, так что её члены кормятся разными способами и с разной скоростью, и социальная информация от них дезориентирует. В подобной ситуации МАД, ведущий к ассортативности образования стай, выгоден всём экотипам [31]. Действительно, проведённые в прошлом полевые опыты по проигрыванию стаям клестов позывов симпатрических экотипов показали, что они оседали тем вероятней, чем меньше были различия между формой их собственных клювов и клювов тех экотипов, чей крик звучал [30].
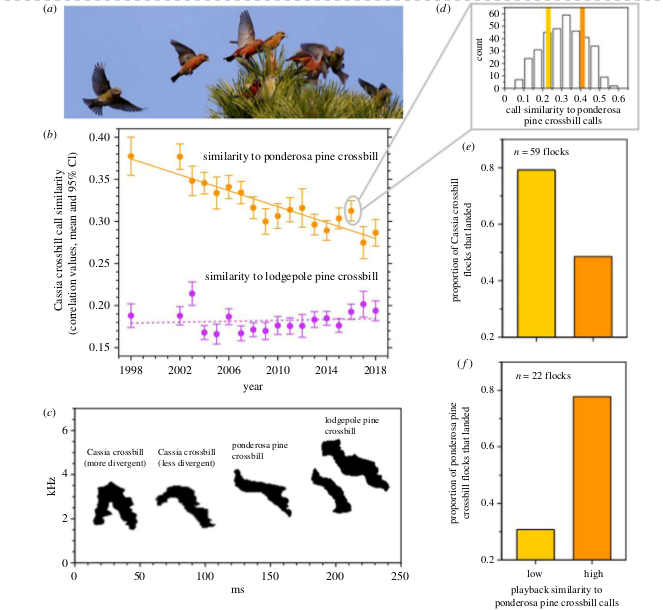 Рисунок 2. Различия контактных позывов клестов Южных холмов (в перечне «племён» №9) и клестов экотипа, специализированного на жёлтой сосне (№8), выросли за последние 20 лет, усилив ассортативность объединения в стаи.
Рисунок 2. Различия контактных позывов клестов Южных холмов (в перечне «племён» №9) и клестов экотипа, специализированного на жёлтой сосне (№8), выросли за последние 20 лет, усилив ассортативность объединения в стаи.
(а) Стая клестов села на вершину сосны. Шесть красно–оранжевых птиц — самцы, четыре желто-зеленые — самки.
(b) Контактные позывы клестов Южных холмов за 20 лет (1998-2018 гг.) дивергировали от позывов клестов экотипа сосны жёлтой (сходство их акустической формы уменьшилось, сплошная линия — регрессия по методу наименьших квадратов; см. текст). Дивергенции от позывов клестов экотипа сосны скрученной за эти же годы не отмечено. Ежегодно наблюдали от 67 до 432 кассиевых клестов, в среднем 247. (c) Спектрограммы позывов клестов разных экотипов, слева направо: кассиевы клесты (с более и менее дивергировавшей акустической формой), клесты экотипов сосны желтой и сосны скрученной. Позывы кассиевых клестов с сильней дивергировавшей акустической формой (крайний слева на спектрограмме) характеризуются большей частотной модуляцией — большим контрастом перехода от восходящих элементов к нисходящим, т.е. более острым «углом» между ними. Сходство её акустической формы с таковой клестов экотипа сосны скрученной всего 0,275, тогда как позывов кассиевых клестов с меньшей частотной модуляцией 0,325.
(d) Распределение позывов, записанных от кассиевых клестов в 2016 г. (в овале, взятом из b), по общему сходству акустической формы с позывом клестов экотипа сосны жёлтой. Ось х — сходство, у — число вокализаций с таким сходством, жёлтая и оранжевая полосы — нижний и верхний квартили распределения.
Примечание. Обратите внимание на значимо большую изменчивость позывов кассиевых клестов по сходству с позывами клестов сосны жёлтой, чем по сходству с позывами клестов сосны скрученной. Рост степени дивергенции позывов за годы наблюдений (b), вызовет большую ассортативность группирования в стаи, поскольку (e) кассиевы клесты с большей вероятностью осаживались проигрыванием позывов, сильней отличающимся от таковых клестов сосны жёлтой, т.е. соответствующих левой, не правой спектрограмме в (с). И наоборот (f) клесты экотипа сосны жёлтой с меньшей вероятностью осаживались проигрыванием позывов кассиевых клестов, менее сходных с их собственными, т. е., с большей частотной модуляцией (таблица 1).
Поскольку клесты выбирают партнеров внутри стаи [29], репродуктивная изоляция между экотипами возникает как побочный продукт ассортативного группирования в стаи, т. е. работой МВД, основанной на сходстве акустических форм контактных криков, коррелятивно связанного с размером клюва [30]. Следовательно, механизмы воспроизводства и дивергенции акустической формы контактных криков, способы их влияния на ассортативность группирования в стаи птиц разных экотипов — ключ к пониманию влияний культурной эволюции на видообразование.
Как и другие вьюрки подсемейства щеглиных Carduelinae, [молодые] клесты выучивают контактные крики родителей и могут видоизменять их всю жизнь [32-34]. Ранее было показано, что кассиевы клесты (Loxia sinesciuris, первоначально описанные как экотип №9 еловиков Северной Америки, но сейчас выделенные в отдельный вид [35]), на протяжении жизни меняют акустическую структуру вокализаций в сторону всё больших отличий от криков генеалогически самого близкого к кассиевому клестому и симпатричного с ним экотипа №2 [28], специализированного к жёлтой сосне [32]. В точности механизм этих прижизненных изменений позывов, увы, неизвестен, но само изменение, скорее всего — приспособительная реакция на значимое ухудшение социальной информации [о качестве кормовых ресурсов и предпочтительных кормовых методах] в смешанных стаях (электронный допматериал, рисунок S1), а это снижает скорость потребления корма [31].
Такой механизм [поддержания ассортативности образования стай] наиболее вероятен: поведение клестов крайне чувствительно к изменениям скорости поступления корма [это монофаг, часто размножающийся зимой, т. е. «напряжение жизни» = интенсивность борьбы за существование здесь значительно выше, чем у прочих вьюрковых] [36], а присоединение клестов к стаям или пребывание там происходит дифференцировано в зависимости от разницы в величине клюва между птицами в стае и присоединяющимися [30]. Аналогичные процессы [аффилиации-дифференциации] также отмечены в других сообществах, где поведение особей изменяется, чтобы взаимодействовать лишь со «своими» и избегать «чужих», когда (или если) они столкнулись с неадаптивными последствиями второго [37-39]. Поскольку крики кассиевых клестов последовательно дивергируют от криков клестов жёлтой сосны, новые поколения кассиевых клестов выучивают всё более непохожие позывы, направленно изменяя популяционную норму вокализаций [30,32]. Действительно, «чужие» позывы выучиваются крайне редко (3 случая из 844 птиц, просмотренных за год, 0,36%, эл. допматериал рис. S2). Поэтому можно ждать дальнейшего роста несходства вокализаций этих двух форм, никак не конвергенции.
Рабочая гипотеза о росте степени дивергенции позывов обоих форм клестов проверена в данной работе. На протяжении 20 лет авторы анализировали аудиозаписи криков клестов, обитающих в Южных Холмах за 20-летний период. Южные Холмы — один из двух небольших горных хребтов на юге штата Айдахо, где обитает оседлая эндемичная форма — кассиев клест (рис. 1; [27,40,41]). Множество разнородных данных (палеоклиматических и палеоботанических реконструкциях [42,43], геномного секвенирования лесов [41]), кассиевы клесты произошли от еловиков широко распространенного экотипа сосны скрученной широкохвойной Pinus contorta latifolia (тип вокализаций и номер «племени» 5 [28]). Леса из этой сосны распространились на обоих хребтах около 5000 лет назад [40,41]. Кассиев клест обособился на Южных Холмах и прилегающем к ним хребте Альбион благодаря отсутствию здесь красной белки (Tamiasciurus hudsonicus). Многочисленная в сосняках из сосны скрученной на крупнейших хребтах Скалистых гор [27,40], она не смогла заселить эти хребты из-за разделяющего их обширного безлесного пространства. диапазоны.
В отсутствие главнейшего конкурента в питании семенами кассиев клест начал коэволюцию с сосной скрученной. У птиц увеличивался клюв, у сосен появлялись всё более прочные шишки, самые трудно открываемые для клестов, чем где-то ещё в обширном ареале этой сосны [27,40]. Этот сценарий недавней дивергенции форм на основе чисто местных различий вместе с широким распространением сосны скрученной поблизости в Скалистых горах ясно показывает, что клесты на кочевках (еловик ежегодно перемещается на сотни километров в поисках шишек) регулярно бывали в регионе на протяжении всей эволюционной истории кассиевых клестов.
Напротив, клесты экотипа сосны жёлтой, видимо, появились на Южных холмах только недавно. Они могли обособиться от птиц экотипа сосны скрученной, чтобы специализироваться на сосне желтой Скалистых гор (Pinus ponderosa scopulorum), когда этот вид в последние 6000 лет расселился на север и восток от Южных холмов с небольшой территории юго-запада США [42]. Отбор поддержит дивергенцию вокализаций кассиевых клестов от позывов обоих экотипов, если это способствует ассортативности группирования в стаи [30], поскольку клесты более мелкоклювых форм, приспособленные к шишками сосны жёлтой и скрученной «своих» районов, неэффективно добывают семена из шишек местной сосны скрученной (единственное хвойное, на котором кормятся все 3 появляющиеся здесь формы [27,40]).
Это снижает качество социальной информации, поступающей к членам смешанных стай, ухудшая кормодобывание кассиевых клестов, буде включаются в смешанные стаи с другими формами (электронный допматериал, рис. S1; [31]). Однако, поскольку кассиевы клесты меньше времени сообитали с клестами экотипа жёлтой сосны и дольше — с клестами экотипа сосны скрученной, их позывы, скорее всего менее дивергировали от первых, чем от вторых и с большей вероятностью будут дивергировать от них, пока не дойдут до состояния равновесия, тогда как с клестами экотипа сосны скрученной они уже скорее всего находятся в этом состоянии.
Помимо проверки рабочей гипотезы о дивергенции акустической структуры вокализаций кассиевых клестов за годы наблюдений, авторы поставили опыты с воспроизведением контактных криков для исследование влияний произошедшей дивергенции вокализаций на поведение симпатрических экотипов в стаях. Также оценивали сходство акустических форм позывов нескольких сим- и аллопатрических пар экотипов клестов, для лучшей интерпретации сравнений позывов кассиевых клестов с клестами двух названных экотипов.
Статья проливает новый свет на роль культурной эволюции [но фактически только на МАД] в [данном случае] симпатрического видообразования, полученные результаты позволяют предположить, что данный механизм был главной действующей силой в обособлении разнотипных форм у клестов.
2. Материалы и методы
(а) Запись вокализаций, подготовка и анализ аудиозаписей
Клестов ловили паутинными сетями, нумерованными службой рыбы и дичи США. Затем их выпускали на волю, сделав аудиозапись контактных криков. Аудиозаписи делали направленным микрофоном ME-66 (Sennheiser Electronic Corporation, Олд-Лайм, Коннектикут) на аналоговом кассетном магнитофоне PMD-222 (1998-2009 гг.) или цифровом магнитофоне PMD-660 (Marantz America, Inc., Аврора, Иллинойс) в 2010-2018 гг. Записи до 2010 г. оцифрованы с частотой дискретизации 44 100 программой Audacity версии 2.2.1. В последующем анализе использовали только те записи, которые однозначно определены как позывы кассиевых клестов. Все записи хранили в виде 16-битных волновых файлов с частотой дискретизации 44 100 Гц.
Спектрограммы записей сделаны программой RAVEN PRO, v. 1.5 [44]. Они визуализированы и проанализированы с использованием окна Ханна, размера быстрого преобразования Фурье в 512 точек и перекрытия окон на 50% на каждом шаге. Каждую запись проверяли на слух и визуально в поиске позывов клестов, не сокрытых фоновым шумом; при их обнаружении, запись сохраняли в виде отдельном файле и использовали в дальнейшем анализе. Затем записи пропускали через полосовой фильтр Ravenpro, чтобы удалить все шумы выше 7500 Гц и ниже 900 Гц.
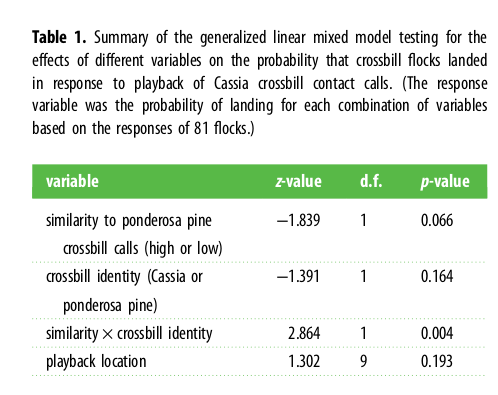
Таблица 1. Краткое описание обобщенной линейной смешанной модели (регрессионный анализ), проверяющей влияние разных переменных на отклик — вероятность оседания стай клестов разных форм после проигрывания контактных криков кассиевых клестов. (Всего тестировали 81 стаю, оценивая шанс приземления при разных комбинациях переменных.)
Обозначения. Столбцы слева направо : переменные, критерий знаков z, степени свободы df, значимость данного фактора р. Строки сверху вниз: сходство акустической формы позывов с таковой у клестов жёлтой сосны, высокая или низкая; принадлежность осаживаемой стаи, кассиевы клесты или клесты жёлтой сосны; взаимодействие двух предыдущих факторов; положение динамика.
 Используя все методы, что описаны выше, авторы делали по 16 записей позывов клестов экотипов сосны жёлтой и скрученной. Их записали на большей части их общего ареала (Вайоминг, Колорадо, Юта и Южные холмы Айдахо) в 1997-2017 гг., поскольку географически их контактные позывы не варьируют [28,45]. Точней, нет географических различий в основном ареале клестов сосны жёлтой (Межгорный Запад), но птицы в Аппалачах и на северо-востоке Соединенных Штатов часто издают отличающийся вариант крика [45]. Однако этот крик никогда не отмечен на Южных холмах и редок в Межгорном Западе, поэтому не включён в анализ.
Используя все методы, что описаны выше, авторы делали по 16 записей позывов клестов экотипов сосны жёлтой и скрученной. Их записали на большей части их общего ареала (Вайоминг, Колорадо, Юта и Южные холмы Айдахо) в 1997-2017 гг., поскольку географически их контактные позывы не варьируют [28,45]. Точней, нет географических различий в основном ареале клестов сосны жёлтой (Межгорный Запад), но птицы в Аппалачах и на северо-востоке Соединенных Штатов часто издают отличающийся вариант крика [45]. Однако этот крик никогда не отмечен на Южных холмах и редок в Межгорном Западе, поэтому не включён в анализ.
Подавляющее большинство клестов этих двух экотипов, пойманных на Южных холмах, там не гнездится [46]; на Южных холмах они появляются в основном в период подвижек (конец весны/начало лета), когда клесты той и другой формы расселяются на большие расстояния, и редки вне этого сезона [40]. Следовательно, эти клесты регулярно приходят из других районов Межгорного запада в Южные холмы и обратно, но немногие из них остаются там круглый год [46]. Таким образом, записи позывов с Межгорного Запада должны фиксировать различия в криках клестов обоих экотипов, с которым сообитают кассиевы клесты. Далее по записям голосов рассчитали среднее попарное сходство каждого позыва кассиева клеста с 16 записями контактных криков клестов двух других экотипов, используя рассчет корреляций в RAVEN PRO [44].
Этот инструмент рассчитывает двумерные кросс-корреляции между двумя аудиовходами для рядов изменений временных и частотных характеристик [звуковых волн]. Мера сходства двух сигналов — максимум корреляции между двумя аудиовходами (ее значения варьируются от 0 до 1, где 1 = идентичность сигналов; доп. материалы, рис. S3; [44]). Кросс-корреляции в RAVENPRO рассчитывали по спектрограммам, они были смещены и нормализованы (в вычислениях игнорировали различия в амплитуде), были извлечены значения линейной мощности, а значения спектрограммы не были искажены.
Кросс-корреляционный анализ лучше всего подходит для сравнения структурно простых и сходных аудиосигналов, таких как контактные позывы разных форм клестов [44]. К тому же результаты кросс-корреляционного анализа обычно сходные с его альтернативами, например, многомерным анализом множества акустических переменных, однако за ним преимущество прямого сравнения общего сходства аудиосигналов, которое может включать неизвестные нам, но значимые для различения сигналов аспекты акустической формы, которые из-за этого не будут выделены и измерены [47,48].
Методы, использованные для оценки степени дивергенции позывов кассиевых клестов от клестов сосны жёлтой, с одной стороны, скрученной с другой, предполагают что позывы двух последних форм относительно кассиевых клестов не менялись за годы наблюдений. Эту гипотезу проверяли, рассчитав сходство между позывами клестов данных двух экотипов, записанных в разные годы, с позывами кассиевых клестов, записанных в эти же годы. Позывы клестов данных двух экотипов, записанные с 1998 по 2018 год на западе США, загружали из Xeno-canto, онлайн-базы данных записей вокализации птиц. Используя вышеописанную кросс-корреляцию, для каждого года рассчитывали близость акустической формы позывов кассиевых клестов с таковой клестов экотипов №2 и №5, записанными в эти же годы. Такие сравнения производили лишь в годы, когда было хотя бы 4 встречи клестов сосны желтой (в среднем 16/год; диапазон = 4-45) или клестов сосны скрученной (12/год; 5-34).
Чтобы определить масштаб долговременных изменений позывов кассиевых клестов [с точки зрения сходства с позывкой клестов экотипов №2 или №5], авторы анализировали вокализации, записанные от данных особей более года подряд. Далее сравнивали общее сходство, основанное на расчёте кросс- корреляций, между позывами тех и других, в первый год, когда их записывали, с аналогичной кросс-корреляцией в последний год наблюдений этих же кассиевых клестов, чтобы определить знак и масштаб изменения вокализаций за соответствующий период времени (в среднем 1,93 года; диапазон: 1-8 лет).
Всех записи подвергали автоматическому спектральному анализу при помощи программы SOUND ANALYSIS PRO 2011 [49] с настройками по умолчанию для зебровых амадин (Taeniopygia guttata). Так исследовали [наличие и выраженность] систематических изменений в [структуре вокализаций:] длительности слогов, совершенству питчей (pitch goodness, мера сочетанности гармоник), средней частоте, частотной и амплитудной модуляциям, энтропии (мера ширины и однородности спектра мощности) и воспринимаемого на слух тона. Записи делили по амплитуде [на фрагменты с минимальным прерыванием звука на 7 мс; слог считался завершённым, когда прерывание звука превышало 10 мс. Судя по измерениям характеристик спектра, долговременные изменения в позывах кассиевых клестов затронули лишь частотную модуляцию, и изменения шли в сторону уменьшения сходства с позывами экотипа жёлтой сосны (рис.2); не отмечено изменений никаких признаков акустической формы сигналов кассиевых коестов, коррелирующих с их сходством с позывами экотипа сосны скрученной.
[Прим. Vinnie sanders oтносительно термина pitch goodness, совершенство питчей. «Я затрудняюсь сказать, как это правильно переводится, т.к. не исключено, что термин «придуман» для специфической области спектрального анализа. Своего рода «добротность тона» — вот только «добротность» вполне конкретный термин, определяющий ширину резонанса, и на англ. обычно пишется как quality factor.
Судя по всему, здесь ищут способом кепстрального анализа периодически повторяющиеся звуки. Есть такой специфический фильтр для ПО спектрального анализа звука как «goodness of pitch» — пики кепстра, вычисленного для гармонических тонов (т.е. сигналов вида основная частота с гармониками. Кепстр же — обратное преобразование Фурье логарифма амплитуды спектра. Своего рода «спектр спектра«.
Умеет такое делать, например, программа Sound Analysis Pro, как раз и использованная авторами. В руководстве по ней приводится картинка, демонстрирующая, как выглядит полученная визуализация «goodness of pitch«: красными стрелками обозначены гармонические звуки (т.е. имеющие выраженные гармоники (повторы сигнала на кратных основной частоте частотах) а синей — негармонический звук (т.е. с “повторами” не на кратных частотах)
«Goodness of pitch» — «добротность (???) высоты звука» или «добротность тона» позволяет выявлять повторы сигналов, как модулированных так и не модулированных. Как чистые тона (без гармоник) так и шумы на таком «спектре спектра» будут давать низкие значения. Оценка периодичности сигнала позволяет выделять вокализованный речевой сигнал. А также есть метод оценки основного тона с помощью кепстрального анализа. Предполагаю, здесь исследователи делают примерно то же, но для птичьего пения].
(б) Сравнительный анализ криков клестов разных экотипов
Здесь авторы сравнивали высоко симпатичные форм клестов-еловиков (одна сосуществует с другой на большей части видового ареала): (i) птиц экотипов сосны жёлтой и скрученной, сообитающих на большей части Межгорного Запада (рис.1), (ii) птиц экотипов западной тсуги и псевдотсуги Мензиса (типы позывов 3 и 4, [29] соответственно), (iii) западной тсуги и ситхинской ели (тип позывов №10; [50]) (все они обитают совместно на большей части Тихоокеанского северо-запада США и Канады, он же Каскадия); (iv) Формы, специализированные на шишках псевдотсуги и сосны скрученной, обитающие совместно на значительной части Межгорного Запада, также как и (v) клесты экотипов псевдотсуги и сосны жёлтой.
Также сравнили вокализации слабо симпатичных форм клестов: (i) экотип сосны жёлтой vs аппалачские клесты (в перечне «племён», т. е. типов контактных криков №1), спорадически сообитающих в небольшом числе по всей Аппалачии и вообще северо-востоку США, (ii) клестов экотипа сосны жёлтой с клестами Сьерра-Мадре (в перечне «племён» №6 [28]), симпатричных на небольшой территории юго-запада США и севера Мексики (iii) экотип сосны скрученной и Сьерра-Мадре, порой совместных на юго-западе США. Географическое распределение их всех описано в Benkman & Young [45]. В данных сравнениях использовали тот же кросс-корреляционный анализ, со следующей величиной выборки по типам позывов (в скобках): тип 1 (15), тип 2 (16), тип 3 (20), тип 4 (17), тип 5 (16), тип 6 (13) и тип 10 (17).
(c) Эксперименты с воспроизведением позывов
Клестам разных форм проигрывали два варианта записей позывов кассиевых клестов — (1) наиболее и (2) наименее сходных с позывом клестов сосны жёлтой. (1) создали случайным выбором 10 записей из всех примерно соответствующих верхнему квартилю распределения пиковых значений корреляции (0,40) 2016 год (средняя значение пиковой корреляции для высоко сходных записей 0,41). В (2) также выбрали 10 записей, но соответствующих нижнему квартилю распределения пиковых значений корреляции (0,23) в 2016 году (рисунок 1d; средняя пиковых корреляций у записей с низким сходством = 0,23).
В каждую группу записи отбирали так, чтобы единственным отличием вокализаций было большее или меньшее сходство с «перекличкой» клестов жёлтой сосны. Такой дизайн дает проверить, влияет ли долговременное снижение сходства криков кассиевых клестов и клестов жёлтой сосны пондерозы на реакцию тестируемых клестов в опытах. После выбора записей для опытов в RAVENPRO создали синтетические записи: один и тот же позыв «размножили» так, что имитировалась временная структура обычных криков клестов (серия из 30 с крика с примерно 88 вокализациями, затем 20 с пауза; см. [30]). Все записи стандартизировали по амплитуде.
Синтетические записи проигрывали свободноживущим клестам с 22 мая по 3 июня 2018 г. через портативный динамик на небольших изолированных участках леса из сосны скрученной (единственное местное хвойное, на котором кормятся клесты). Позывы транслировали в 10 точках, расположенных не менее 500 м друг от друга, утром и вечером портативным усилителем Pignose 7–100 Legendary (Pignose-Gorilla, Лас-Вегас, Невада). В каждой точке проигрывали записи и (1), и (2); порядок воспроизведения групп записей меняли каждый день так, чтобы (1) и (2) воспроизвелись с одной и утром и вечером.
Все опыты по проигрыванию провёл один и тот же человек (C.K.P.), не знающий какая запись (1), какая (2). За 120-мин каждого проигрывания авторы фиксировали, приземляются ли пролетающие над ними клесты на сосны рядом с громкоговорителем или летят дальше. Данный эксперимент имитирует поведение стай клестов в природе: они громко зовут пролетающих над ними других клестов; это чрезвычайно распространенное поведение, стимулирующее формирование и укрупнение стай [надо думать, в наиболее обильных кормом местах, где птицы задерживаются подольше] [29,30]. Поэтому регистрируя, приземлятся ли стаи в ответ на воспроизведение, можно определить готовность клестов к объединению в одну стала сородичами, различающимися акустической формой позывов [30].
Громкоговоритель поставили установлен на краю лесного участка на высоте примерно 1,5 м. Формы клестов определяли на слух, по характерным контактным крикам; кассиевы клесты и клесты жёлтой сосны преобладали среди клестов, отмеченных на Южных холмах в 2018 году, и только они присутствовали при проигрывании. Так протестировали 81 стаю, все чистые, не смешанные. Как и в предыдущем исследовании [30], каждую стаю считали независимым испытанием, т. е. n=81.
(d) Статобработка данных
Её проводили в R [51], использовали лишь двусторонние тесты, «усы» на рисунках — 95%-ные доверительные интервалы. Никакие переменные не были изменены перед анализом, поскольку были соблюдены предположения о нормальности. Для оценки долговременных изменений сходства криков и частотной модуляции в популяции кассиевых клестов использованы среднегодовые значения линейной регрессии, также использованные оценке взаимосвязи степени сходства [с криком клестов жёлтой сосны] и выраженностью частотной модуляции. Парные t-тесты использовали для сравнения сходства криков повторно отловленных через несколько лет [меченых] кассиевых клестов с криками экотипов сосен жёлтой и скрученной.
Данные опытов по проигрыванию анализировали с помощью биномиальной обобщенной линейной смешанной модели, а также применяли logit link function для двоичных исходов R [51], пакет «lme4» [52]. Факторами, чье влияние исследовали, было сходство проигрываемых криков с таковыми экотипа жёлтой сосны, принадлежность осаживаемой стаи еловиков (кассиевы клесты или клесты жёлтой сосны) Зависело от того, приземлялись ли стаи скрещенных клювов в ответ на воспроизведение (да или нет), откликом — приземляется стая или нет, случайный эффект — место проигрывания.
3. Результаты
Анализ стандартизированных записей контактных криков 3242 кассиевых клестов Южных Холмов в 1998-2018 гг., показывает значимое смещение признаков акустической структуры вокализаций.
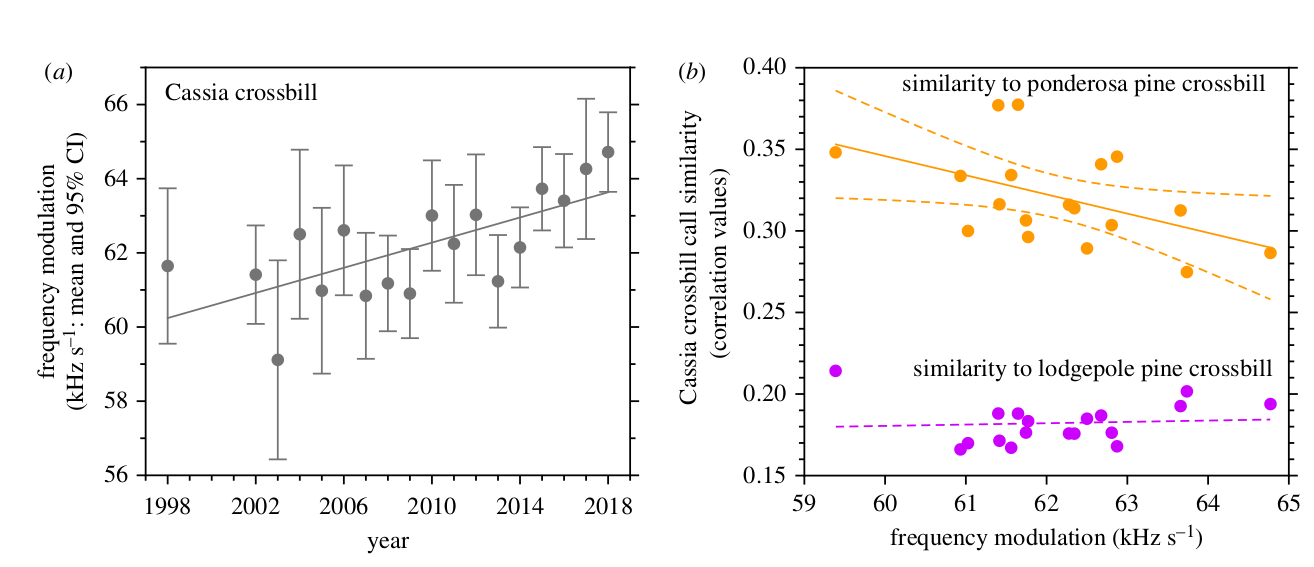 Рисунок 3. Наблюдаемые изменения сходства криков, показанные на рисунке 2b, видимо, результат увеличения частотной модуляции позывов кассиевых клестов с 1998 по 2018 год (а). Именно усиление частотной модуляции уменьшает сходство позывов кассиевых клестов с клестами экотипа сосны жёлтой, не влияя на сходство с клестами сосны скрученной (b); каждое из этих изменений связано с увеличением частоты модуляции. Каждая точка — отдельный год, как показано в (а).
Рисунок 3. Наблюдаемые изменения сходства криков, показанные на рисунке 2b, видимо, результат увеличения частотной модуляции позывов кассиевых клестов с 1998 по 2018 год (а). Именно усиление частотной модуляции уменьшает сходство позывов кассиевых клестов с клестами экотипа сосны жёлтой, не влияя на сходство с клестами сосны скрученной (b); каждое из этих изменений связано с увеличением частоты модуляции. Каждая точка — отдельный год, как показано в (а).
Мера сходства акустической формы позывов, кросскорреляция, неуклонно снижалось от 0,377 в до 0,287 в 2018 году (уменьшение на 24% за 20 лет, рис.2b; аппроксимация линейной регрессией среднее сходство = 9,8248-0,0047*год); r2= 0,84, df = 16, p < 0,0001). Сходный результат дал анализ учитывавший потенциальные изменения числа позывов клестов сосны жёлтой, записанных в разные годы (среднее сходство = 9,9810 − 0,0048 * год; r2= 0,70, df = 7, p = 0,009), включая почти одинаковую скорость снижения сходства за данные 20 лет (0,0047 год−1 в в первом анализе и 0,0048 год−1 во втором анализе). На популяционном уровне данная дивергенция, видимо, создана долговременным ростом частотной модуляции криков кассиевых клестов (рис.2с и рис.3a; частотная модуляция = -234,8726 + 0,1478 * год; r2= 0,47, df=16, p=0,002), поскольку именно с ним отрицательно коррелирует сходство их акустических форм с формой позывов клестов сосны жёлтой (рис.3b; сходство позывов двух форм клестов = 1,0510–0,0118 * частотная модуляция в позывах кассиевых клестов; r2 = 0,47, df=16, p=0,002).
= 0,24, коэффициент полезного действия = 16, р = 0,038).
Из 6 прочих измеренных автором характеристик акустической формы позывов кассиевых клестов не изменялась за годы наблюдений таким образом, чтобы обуславливать зафиксированные изменения значений степени сходства с клестами сосны жёлтой. В противоположность направленному росту несходства с позывами клестов сосновы жёлтой, сходство с клестами сосны скрученной не изменилось значимо за тот же период (рис. 2b; r2= 0,01, df = 16, p = 0,65), даже с учетом потенциальных изменений числа встреч и записей этой формы со временем (r2=0,06, df = 5, p=0,64). Такое отсутствие дивергенции вполне ожидаемо, учитывая уже имеющиеся значительные различия между ними и кассиевых клестами (рис. 2b), возможно, вследствие их долговременной симпатрии.
Кроме того, степень сходства позывов клестов сосны скрученной и кассиевых клестов не связано с частотной модуляцией криков тех и других (рис.3b; сходство криков клестов сосны скрученной = 0,1314 + 0,0008 * частотная модуляция; r2= 0,01, d.f. = 16, p = 0,76). Отсюда нельзя ждать дальнейшего расхождения (или сближения) с ними позывов кассиевых клестов как побочного следствия дивергенции их от позывов клестов сосны желтой.
Названные закономерности популяционного уровня проявляются в сонаправленных изменениях позывов отдельных птиц в эти же годы. Крики 801 кассиевых клестов, отмечавшихся нескольку лет, со временем становились всё больше и больше отличными от криков клестов сосны жёлтой (парный t-критерий: t779=5,25, p < 0,0001; примеры индивидуальных изменений со временем см. электронный доп.материал, рис.S4), что подтверждает предыдущие выводы, основанные на на выборке всего из 114 особей [32]. Среднее уменьшение сходства у отдельных особей за год (0,0123 ± 0,0031, средняя± s.e.) в 2,6 раза больше среднего снижения за год на уровне популяции (0,0047 ± 0,0005).
Это понятно, ибо потомство не может в совершенстве имитировать крики родителей [32,34], а несовершенная передача изменений между поколениями снижает несходство на уровне популяции сравнительно с индивидуальным. Сходство позывов отдельных особей кассиевых клестов, прослеженных несколько лет, с позывами клестов сосны скрученной, значимо не изменилось, подтвердив тенденцию популяционного уровня (парный t-критерий: t779= -0,04, p = 0,97).
Предположив недавнее установление симпатрии между клестами кассии и сосны желтой, [41-43], авторы предсказали, что сходство их позывов будет примерно на уровне такового между парами аллопатрических экотипов клестов, предположительно, еще не испытывали смещения признаков их акустической формы. Действительно, первоначальное сходство позывов кассиевых клестов и клестов сосны жёлтой в 1998 году (0,377) сравнимо со сходством окликов между тремя преимущественно аллопатрическими парами экотипов (средняя 0,317; диапазон: 0.161–0.403; электронный доп.материал, рисунок S5).
Напротив, уровень (не)сходства позывов кассиевых клестов и клестов сосны скрученной за последние 20 лет (0,166–0,214) ближе к (не)сходству криков между шестью высокосимпричными парами экотипов еловиков (средняя: 0,259; диапазон: 0.206–0.350; электронный дополнительный материал, рис.S6). Вероятно, долгая симпатии между ними стала причиной столь значительной дивергенции.
Ключевой прогноз рабочей гипотезы про смещение признаков, что оно уменьшает риск неадаптивного образования стай из птиц разных форм [30]; поскольку клесты выбирают партнеров в своей стае [29], оно повысит ассортативность спаривания внутри каждой формы [30]. В соответствии с ним, чем сильнее росло несходство криков кассиевых клестов и клестов сосны жёлтой, тем реже вторые осаживались проигрыванием позывов первых (сдвиг от высокого к низкому сходству сигналов на рисунке 2ef). Иными словами, существует обратная связь между сходством криков двух формами и рисками появления гетероспецифических стай (р = 0,004; таблица 1). Последние сильно уменьшились всего за 20 лет за счёт дивергенции акустической формы сигналов (рис.2b), ассортативное спаривание -побочное следствие всего этого [30].
4. Обсуждение
Полученные результаты показывают, что у названных форм клестов выучивание позывов родителей вместе с возможностью модификации их на индивидуальном уровне ведут к дивергенции, а не конвергенции их вокализаций (рис.2b). Что согласуется с прочими данными: палеоботаническими [42,43], генетическими и климатическими реконструкции лесов [41], сравнением акустических форм позывов сим- и аллопатрических форм клестов, указывающими на недавнее появление симпатрии между ними, в сравнении с таковой между кассиевыми клестами и клестами сосны скрученной. Это, по-видимому, объясняет, почему на коротких временных интервалах дивергенция в первом случае происходит, во втором нет (рис.2b).
Эксперименты с воспроизведением криков показывают, что выявленная дивергенция за эти же 20 лет снижает риск изменения гетероспецифического объединения обоих форм коестов в смешанных стаях (рис.2ef; таблица 1). Очевидно, это усилит ассортативность спаривания в каждой форме, тем самым способствуя видообразованию, поскольку клесты выбирают партнеров из стаи [29,30]. Данный вывод имеет косвенное подтверждение: в Скалистых горах поведенческая изоляция [9] между клестами экотипов сосны жёлтой и скрученной усиливалась с ростом степени ассортативности их объединения в стаи (r2= 0,72, p = 0,033, n = 6 сезонов размножения и 254 возникшие пары, C.K.Porter, 2019, неопубл.данные).
Если бы наблюдавшаяся с 1998 по 2018 год дивергенция позывов кассиевых клестов и клестов сосны жёлтой наблюдалась и раньше (рис. 2b) это значило бы ее начало менее ста лет назад (электронный доп.материал). Однако, смещение признаков будет быстрым лишь при промежуточном уровне сходства позывов, особенно при низкой наследуемости [53] имеющей место быть у кассиева клеста [32]. Поэтому отмеченная авторами быстрая дивергенция может быть очень недавней (рис.2b); ей предшествовал период много более медленной дивергенции на ранних стадиях симпатрии, при значительно большие сходстве формы их позывов.
Как долго это продолжалось, пока неизвестно, но степень недавнего расхождения, измеренная в данной работе, указывает на то, что регулярная симпатрия на большой территории клестов сосны жёлтой и кассиевых может быть даже более поздним явлением, чем авторы предполагают сейчас.
В отличие от большинства случаев экологического видообразования, здесь рост приспособленности от дивергенции сигналов создан не пресечением межвидового скрещивания (вследствие смещения признаков; [54]), но уменьшением риска формирования смешанных стай разных, кормление в которых менее эффективно (рис. 2еf; таблица 1), чем в стан одной формы. «Свои» сородичи создают друг для друга более качественную социальную информацию о запасах шишек, чем «чужие» (электронные вспомогательные материалы, рис. S1), благодаря чему скорость поступления пищи в первом случае значимо выше, чем во втором [31].
Смешанные стаи могут быть вредны ещё и потому, что клесты, лишённые пищи, вырабатывают гормоны стресса только в том случае, если сочлены стаи также лишены [55]. Поэтому в такой стае особи, питающиеся хуже всего, будут стрессировать всех остальных; они же больше подвержены стрессу в смешанной стае, чем в «чистой». Кроме того, частота межвидового спаривания между кассиевыми клестами и клестами жёлтой сосны все эти годы была очень низка (0,7% из 1704 гнездящихся пар с 2001 по 2007 год в Саут-Хиллз; [40,56]).
Вместе с предположительно сниженной на 12% приспособленностью гибридов между этими таксонами [57] это привело бы к слабому естественному отбору против них, которого было бы недостаточно для объяснения наблюдаемой дивергенции позывов (рис. 2b). Отсюда предположение, что именно преимущества присоединения к «своим», (электронный дополнительный материал, рисунок S1), отнюдь не отбор против гибридов или смешанных пар, основной адаптивный механизм, определяющим недавнюю эволюцию позывов кассиевых клестов.
Как отмечалось выше, в период с 2001 по 2007 год уровень скрещивания между кассиевыми клестами и клестами сосны желтой был достаточно низким [40] даже в те годы, когда их позывы были наиболее похожи (среднее сходство: 0,346), чем в 2018 году, когда сходство уменьшилось (0,287). Следовательно, наблюдавшееся уменьшение сходства вызвано не им (рис.1b) а сокращением риска формирования гетероспецифических стай (рис.2ef). Поэтому последующее уменьшение частоты образования смешанных пар вряд ли будет значительным в абсолютном смысле (т. е. в десятых долях процента, и поэтому их чрезвычайно трудно обнаружить без обширной выборки). Однако это не означает, что даже незначительное сокращение скрещиваний несущественно для видообразования кассиевых клестов: даже низкий темп интрогрессии в системах с почти полной репродуктивной изоляцией может препятствовать видообразованию [58-61].
Кроме того, если выявленная дивергенция позывов увеличит ассортативность скрещивания, это может быть особенно важно: усиление репродуктивной изоляции затруднительно тем больше, чем более она полная. Поэтому все усилители ассортативности, когда репродуктивная изоляция очень сильна, но неполна, в видообразовании могут быть важнее всего.
Выявленное вторами смещение признаков акустической структуры вокализаций у симпатрических форм (рис.2b), возникло вследствие социального обучения, работающего в системе МАД [32], возможно, для понижения риска объединения в одну стаю с особями других форм, снижающего успешность кормления (электронный дополнительный материал, S1). Меняющиеся вокализации родителей выучиваются потомством [32,34], приводя к дивергенции позывов уже на популяционном уровне (рис.2b) и ещё больше снижая риски формирования смешанных стай данных форм (рис.2ef). Здесь контактные крики выступают маркерами принадлежности, опосредующими группирование в стаи и изменяемые культурной эволюцией.
Поскольку клесты круглый год держатся в стаях и в них же выбирают партнеров [29] отмеченный рост препятствий к скрещиванию, возможно, побочный продукт выявленного смещения признаков. Хотя данное исследование первое из известных авторам, обнаружившее всё вышеописанное в природе, данный механизм был предложен как рабочая гипотеза для объяснения формирования симпатрических экотипов кашалотов Physeter macrocephalus [65,66], дельфинов-афалин Tursiops truncatus [66,67], как и формирования генетически полностью обособленных экотипов косаток Orchinus orca [66,68,69]. Отдельные составляющие этого многоступенчатого процесса выявлены у широкого круга видов животных [37-39,70-75].
Поэтому сценарий, отмеченный у клестов-еловиков, может быть важен и в общем случае, для диверсификации на видовом уровне. Сейчас большинство исследований роли культурной эволюции в дивергенции популяций и видообразовании концентрируются на ней как способе возникновения репродуктивной изоляции в аллопатрических популяциях [10-12,14-19,25,26,54]. Авторский результат вместе с другими похожими показывает ещё одну возможность действия культурной эволюции в формообразовании: содействия раз начавшемуся экологическому видообразованию через содействие дифференциации экотипов, движимой дизруптивным отбором, её усиление и доведение до конца за счёт гетерофобии (присоединением к «своим» и отвержением «чужих», со смещением признаков, делящих на тех и других, в зоне симпатрии), созданной механизмами социального обучения, ярчайший из которых — выучивание песен, позывов или кормовых методов при подражании «модели», опосредуемый (видимо, во всех этих случаях) зеркальными нейронами.

Рекомендуем прочесть
 Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки Быстрая перестройка фенологии в новых местах гнездования ласточек
Быстрая перестройка фенологии в новых местах гнездования ласточек Расшифровка сигналов агрессии у коростелей
Расшифровка сигналов агрессии у коростелей Как соотносятся прямое приспособление и эволюционные изменения?
Как соотносятся прямое приспособление и эволюционные изменения? Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла»
Отбор адаптивной и неадаптивной пластичности в экспрессии генов при приспособлении анолисов к городскому «острову тепла» Ещё про «конформизм» в трансляции поведенческих стереотипов у птиц
Ещё про «конформизм» в трансляции поведенческих стереотипов у птиц Клесты-еловики США: дифференциация «племён» и специализация к шишкам хвойных
Клесты-еловики США: дифференциация «племён» и специализация к шишкам хвойных Быстрая эволюция в связи с урбанизацией: примеры
Быстрая эволюция в связи с урбанизацией: примеры
