
Самец и самка чечевичника на кормушке в городе.
В продолжение темы, как соотносятся прямые приспособления и изменения под действием отбора
Резюме. Везде урбанизация идёт много быстрей и «бескровней» (с многажды меньшим уровнем элиминации), чем следует из господствующей сейчас синтетической теории эволюции (СТЭ). От первых поселений в урбосреде до формирования специализированной городской популяции вида занимает ~10-20 лет у видов-потенциальных урбанистов и 30-60 лет у видов, сперва вытесненных из города и “отступавших” к периферии региона, но вернувшихся туда в ходе возвратной урбанизации.
Везде освоение города заняло ~8-13 поколений, что явно недостаточно для распространения адаптивных изменений внутри популяции тем способом, который СТЭ полагает главным или даже единственным — «поиском» генных вариантов, адаптивных при действии городских факторов неблагоприятности, и ответственных за появление всей новизны в морфологии, поведении т. д., что фиксируется натуралистами при урбанизации «диких» видов. Второй важный момент, отмеченный и в рассказе, что не так с синтетической теорией эволюции: что проходят отбор не интактные особи, но уже изменившиеся в борьбе за существования, данные изменения — плод прямого приспособления, следствие «напряжения сил», неотделимого от этой борьбы, на что обращал внимание Томас Роберт Мальтус.
Поэтому (микро)эволюция, сплавленная с экспансией — направленным освоением урбосреды, не «ищет» полезных приспособлений, не «ждёт» их возникновения, а «шьёт из готового». Адаптации, “выработанные” в специализированных городских популяциях, суть амальгама прямого приспособления с (несколько отстающими по времени) изменениями под действием отбора: на уровне генов они, как правило, отстают от сдвигов поведения и морфофизиологии. Отбор закрепляет общий результат того и другого, но не создаёт связанной с ними новизны. Об этом рассказывается на двух примерах: сопряжения генетических и эпигенетических изменений при урбанизации разных видов, связанных с изменением функционирования одних и тех же структур организма, и поиска компромисса между отчасти противоречащими друг другу требованиями к форме клюва и частотной структуре пения при урбанизации мексиканского чечевичника.
Урбанизация «диких» видов противоречит прогнозу СТЭ
Содержание
- 1 Урбанизация «диких» видов противоречит прогнозу СТЭ
- 2 Как микроэволюция «шьёт из готового»
- 3 Поведенческие изменения у «городских» чечевичников «тянут» за собой морфологию
- 4 Урбанизация и структура песни вьюрков сопряжённо меняются вдоль городского градиента
- 5 Сопряжённое варьирование формы клюва и характеристик песни вдоль городского градиента
- 6 Обсуждение
Везде урбанизация идёт много быстрей и «бескровней» (с многажды меньшим уровнем элиминации1), чем следует из господствующей сейчас синтетической теории эволюции (СТЭ). От первых поселений в урбосреде до формирования специализированной городской популяции вида занимает ~10-20 лет у видов-потенциальных урбанистов и 30-60 лет у видов, сперва вытесненных из города и “отступавших” к периферии региона, но вернувшихся туда в ходе возвратной урбанизации. Везде освоение города заняло ~8-13 поколений, что явно недостаточно для распространения адаптивных изменений внутри популяции тем способом, который СТЭ полагает главным или даже единственным — «поиском» генных вариантов, адаптивных при действии городских факторов неблагоприятности, и ответственных за появление всей новизны в морфологии, поведении т. д., что фиксируется натуралистами при урбанизации «диких» видов (Фридман, Ерёмкин, 2009, см. примеры в табл.1).
Таблица 1. Примерная скорость урбанизации «диких» видов птиц в различных регионах
|
Вид, регион, |
Даты начала урбанизации, годы, примерная скорость процесса, лет |
Число сменившихся поколений/ среднеожидаемая продолжительность жизни особей (СОПЖ) |
Источник |
|
Pica pica (Варшава)* |
Начало 1960-х, 10-20 |
4-5/2 |
Luniak et al., 1997 |
|
Pica pica (Прага)* |
1970е, 20 |
10/2 |
Stastny et al., 2005 |
|
Pica pica (Прага)* |
1970е, 30 |
15/2 |
Iankov, 2005 |
|
Turdus pilaris (Варшава) |
1975, 10-15 |
9-10/1,6 |
Luniak, 2004 |
|
Turdus pilaris (Трускавец) |
Конец 1980-х, 10-15 |
9-10/1,6 |
Константинов, 2001 |
|
Turdus pilaris (Харьков) |
1987, 10 |
9-10/1,6 |
Чаплыгина, Кривицкий, 1996 |
|
Turdus merula (Кёнгсберг)* |
1889-1938 гг. |
30/1,5 |
Tischler, 1941. |
|
Accipiter gentilis (Москва) |
1975, 20-25 |
7/3,5 |
Ерёмкин, Очагов, 1998 |
|
Accipiter gentilis (Киев) |
Конец 1980-х, 15-20 |
7/3,5 |
Матус, 2003 |
|
Coccothraustes coccthraustes (Москва) |
Начало 1970-х, 15-20 |
10-11/1,4* |
Ерёмкин, Очагов, 1998 |
|
Aythya fuligula (Москва) |
1975, 20-25 |
12-13/1,9* |
Ерёмкин, 2004 |
|
Asio otus (Москва) |
Конец 1970-х, 25-30 |
9-11/2,9* |
Воронецкий, Леонов, 2003 |
|
Anas platyrhynchos (Москва) |
Конец 1950-х, 15-20 |
14-15/1,4 |
Авилова, 1998 |
|
Anas platyrhynchos (Ленинград) |
Конец 1960-х, 15-20 |
14-15/1,4 |
Храбрый, 1994 |
|
Cyanosylvia svecica (Москва) |
Середина 1970-х, 20 |
8-9/2,3 |
Ерёмкин, 2004 |
|
Carduelis chloris (Москва) |
Середина 1960-х, 15-20 |
10-11/1,8 |
Ерёмкин, 2004; Б.С.Самойлов, личн. сообщ. |
|
Ficedula hypoleuca (Москва) |
Начало 1980-х, 10-15 |
10-11/1,4 |
Ерёмкин, 2004; Б.С.Самойлов, личн. сообщ. |
|
Columba palumbus (Черновцы) |
Середина 1980-х, 10-15 |
6-7/2,2 |
Скильский и др., 1997 |
|
Columba palumbus (Львов) |
Середина 1990-х, 10 |
6-7/2,2 |
Сенник, Хорняк, 2003 |
|
Columba palumbus (Калининград)* |
1974-1989, 19-34 |
8 или 16/2,2 |
Астафьева и др., 2011 |
|
Parus major (Москва) |
Середина 1960-х, 15-20 |
20-23/0,9 |
Ерёмкин, 2004; Б.С.Самойлов, личн. сообщ. |
|
P.caeruleus (Москва) |
Середина 1970-х, 15-20 |
20-23/0,9 |
Ерёмкин, 2004; Б.С.Самойлов, личн. сообщ. |
|
Corvus brachyrhynchos (разные города США) |
1950-1970-е (в зависимости от региона), 10-20 |
9-10/2,2* |
Marzluff et al., 2001a |
|
Corvus corone orientalis (Иркутск) |
1967 или 1972 – 1979, 10 лет |
9-10/2,2* |
Липин и др., 1983 |
Примечания. Скорость урбанизации вида рассчитывали по продолжительности периода от начала регулярного гнездования на территории города до формирования устойчивой и жизнеспособной городской популяции. Данные по СОПЖ взяты из монографии В.А.Паевского (1985), табл.30: число сменившихся поколений считали равным длительности процесса урбанизации / СОПЖ во «внегородских» популяциях вида, приток особей из которых «подпитывает» рост городских популяций. * — использованы данные по СОПЖ близких видов: для C.coccthraustes – данные по C.chloris, для A.fuligula — данные по Bucephala clangula, для Asio otus – по Strix aluco, для С. brachyrhynchos и С.corone orientalis — по европейским популяциям серой вороны.
Источники: Фридман, Ерёмкин, 2009 и # — материалы, предоставленные Е.Г. Лыковым. Вторые показывают не момент становления, а конец формирования городской популяции, т. е. сроки распространения по всей территории города и стабилизацию численности.
Пусть данный процесс идёт, как предполагается в СТЭ: “нужный” вариант сперва возникает в “нужной” популяции или “попадает” туда через гибридизацию, комбинативную изменчивость и пр. внеотборные процессы, а дальше “распространяется” там отбором. Тогда для фиксации единичных приспособительных изменений в подразделённых популяциях с эффективной численностью в первые сотни особей (если исходная частота их невелика), должны смениться десятки-первые сотни поколений (даже при значительности их селективного преимущества). Число поколений, необходимых для распространения полезных изменений в городской популяции, оценивают по формуле для скорости изменения частоты аллеля за поколение ∆q = -spq/(1-q2), где s = коэффициент отбора против рецессивных гетерозигот, p и q — частоты альтернативных аллелей гена, «поддерживаемого» или «элиминируемого» отбором (Айала, Кайгер, 1988, табл.24.5).
Эта мажорирующая2 оценка сделана в предположении полного доминирования отбираемых уклонений, зависимости приспособительно-ценных признаков только от одного гена и 100%-ной наследуемости; на деле поколений (и времени) нужно больше. Ведь большинство приспособлений, фиксируемых в городской популяции, суть поведенческие инновации (вместе с морфофизиологическими изменениями, которые «тянет» за собой нужда в их устойчивой реализации в онтогенезе, см.ниже). Их наследуемость (h2) ~0,2-0,3, что ещё больше увеличивает число поколений, нужное для распространения в популяции единичного адаптивного изменения вышеописанным способом.
Соединение отдельных приспособлений в целостный комплекс, описываемый как «своеобразные черты биологии городских популяций», под действием этого же механизма должно быть ещё более долгим процессом (Айала, Кайгер, 1988) — однако он возникает исключительно быстро и сразу во всех основных чертах, дальше лишь «конкретизирующихся» и развивающихся. (Микро)эволюция, сплавленная с экспансией — направленным освоением урбосреды, не «ищет» полезных приспособлений, не «ждёт» их возникновения, а «шьёт из готового3».
Всё вышесказанное особенно очевидно, когда речь идёт о недавно возникших мелких субпопуляциях, «черпающих» резервы для роста из обширного «резервуара» — региональной популяции многажды большей численности. Практически все городские популяции в период формирования и экспоненциального роста численности, до наступления стабилизации, демографически несамостоятельны, существуют за счёт «подтока» особей из ближних пригородов (Фридман, Ерёмкин, 2009; Фридман, Суслов, 2022, разд.2.3-2.4).
Как микроэволюция «шьёт из готового»
Адаптации, “выработанные” в специализированных городских популяциях, суть амальгама прямого приспособления с (несколько отстающими по времени) изменениями под действием отбора: на уровне генов они, как правило, отстают от сдвигов поведения и морфофизиологии (Фридман, Суслов, 2022, разд.2.5-2.7). Отбор закрепляет общий результат того и другого.
Прямое приспособление фиксируется уже в первом поколении в диапазоне от поведенческих инноваций до эпигенетических изменений — “меток” в виде метилирования генов, присущего особям, вырастающим в экстремальной среде. Фактически это длительные модификации, наиболее известные у растений и беспозвоночных, но на деле присущие всем живым существам без изъятия. Так, депрессия матери в пренатальном периоде меняет паттерн метилирования промотора SERT у плода, а его общий уровень связан с плохим обращением в детстве. Уровни метилирования DRD4 и SERT также отрицательно коррелируют с развитием симптомов синдрома дефицита внимания и гиперактивности. На протяжении жизни у однояйцевых близнецов накапливаются различия в метилировании генов SERT и DRD4, отражая возрастающее несходство их биографий и жизненных обстоятельств (Riyahi et al., 2015).
Как и в случае с человеческим детством, вырастание в экстремальной среде создаёт «отпечаток» в виде метилирования генов, меняющего их экспрессию в сторону, обеспечивающую первый этап адаптации — интенсификацию функции, «попавшей» под действие стрессоров, чтобы и в этих условиях «жить, ничего не меняя». Позже, уже на втором этапе приспособления, они замещаются генетическими изменениями с противоположным эффектом: обуславливая сдвиг поведения (или морфофизиологии вида, или создавая способность к временному, «здесь и сейчас» сдвигу этого поведения и физиологии), они порождают ситуацию, когда ранее экстремальная среда делается если не благоприятной, то пермиссивной — какое-то время её можно эксплуатировать, но потом надо отдыхать в рефугиуме или хотя бы на время уменьшить активность, восстановить силы (Фридман, Суслов, 2022).
Так, отбор «городских» вариантов DRD4 больших синиц сопровождается и эпигенетическими изменениями. Так, изучались птицы из городских парков Барселоны в сравнении с синицами из двух участков природных ландшафтов. В урбопопуляциях уровень метилирования CPG-островов4 в его промоторе и экзоне 3 (содержит два CPG-острова, один из которых заходит в интрон 3 — главную “цель” отбора генов в урбосреде) был значимо, хотя и не сильно, выше в образцах ДНК как из мозга, так и из крови (на ≈1%). CPG-острова промотора данного гена метилированы (8%) относительно экзона 3 DRD4 (93%) равным образом в городских, так и в исходных популяциях.
Промотор SERT беден CPG. Метилирование одного динуклеотида CPG в позиции 288 п.н. от начала транскрипции, коррелирует с интенсивностью исследования у «городских» птиц. Полиморфизм SNP234, расположенный внутри минимального промотора SERT, коррелирует с поведением поиска новизны в неволе. Аллель, усиливающий последнее, чаще встречается у «городских» синиц (Riyahi et al., 2015).
Действительно, метилирование «генов индивидуальности» DRD4 и SERT в «городских» популяциях птиц значимо отличается от такового в исходных и как минимум у больших синиц различия связаны с изменением индивидуальности при урбанизации. У них же различие транскриптомов между популяциями связывают с эпигенетическими изменениями уровней экспрессии генов (Riyahi et al., 2015; Verhulst et al. 2016; Watson et al., 2017).
У дарвиновых вьюрков Geospiza fortis и G.fuliginosa на о.Санта-Крус урбопопуляции отличаются эпигенетически, степенью метилирования разных участков ДНК: морфологические отличия от исходных популяций отмечены лишь у первого вида. Значительность различий, связанных с урбанизацией, подчёркнута называнием их «драматическими». Не найдено различий в изменчивости числа копий генов (англ. copy number variation) между популяциями (McNew et al., 2017).
Обычны случаи, когда в конкретных городских популяциях изменения поведения и/или морфофизиологии уже присутствуют и безусловно наследственны, т.е. развиваются у “городских”, но не “сельских” особей при выращивании в стандартных условиях лаборатории (Фридман, Суслов, 2022, глава 2.5). Отбор на уровне генов здесь ещё не заметен, он появится позже. Поэтому приспособительные изменения генов (особенно, “генов урбанизации”, DRD4 и SERT) отмечены лишь в популяциях, существующих дольше и/или освоивших более крупные города.
В четырёх городских популяциях большой синицы (595 птиц) из Дании в отсеквенированном гене DRD4 найдено 98 однонуклеотидных замен (SNP), ассоциируемых с изменением поискового поведения. Они связаны как с кодирующими, так и с некодирующими участками, первые несут следы позитивного отбора. Наибольшая ассоциация с изменением поисковой активности — с SNP экзона 3 (особенно в 830й позиции), поменьше — с SNP экзонов 2 и 4. Изменения шли параллельно во всех урбопопуляциях, максимум ассоциаций отмечен в городах Вестерхейде и Ловерсмер. Там же, но не в «лесных» поселениях (Boshoek и лес Витам близ Оксфорда, Англия) они связаны и с исследовательским поведением (Müller et al., 2013а).
Вообще, метилирование ДНК
а) различается между городскими и исходными популяциями, включая локусы DRD4 и SERT;
б) как минимум у больших синиц, дарвиновых вьюрков и кроличьих сычей связано с поведенческими различиями между ними;
в) анализ их транскриптомов показывает устойчивые различия в профилях экспрессии множества разных генов между «городом» и исходными биотопами (Riyahi et al. 2015; Verhulst et al. 2016; Watson et al., 2017; McNew et al., 2017). Что говорит о значимости эпигенетических различий для канализации изменений, связанных с их освоением. Последнее согласуется с важностью стресса, особенно в раннем онтогенезе, для программирования изменений, повышающих устойчивость существования в будущей среде обитания (Suslov, 2013; Роговин, 2019).
C другой стороны, урбанизация связана с селективными изменениями множества разных генов одновременно, не только «знаменитых» DRD4 и SERT. Так, полногеномный анализ «городских» vs «сельских» кроличьих сычей Athene cunicalaria (213 птиц из трёх пар местообитаний обоих типов) выявил селективные изменения 98 генов, в основном экспрессируемых в головном мозге и связанных с синапсами и проекциями нейронов. «Городские» популяции также отличаются от «сельских» полиморфизмами в промоторной области гена SERT, важного для последующего рассмотрения (Müller et al., 2020). Видимо, большинство генов, в процессе урбанизации оказывающихся под отбором, отражают частные приспособления к данному конкретному городу, вроде гена RARS1 первой хромосомы гребенчатых анолисов, освоивших города о.Пуэрто-Рико.
Селективные изменения в генах SERT и DRD4 в связи с урбанизацией интересны своей однородностью у птиц, очень разных экологически и филогенетически: варьирует число накопленных изменений, «распространение» в гене позиций, изменённых отбором, однако они идут в одном направлении в географически очень разных, несвязанных друг с другом городских популяциях — скажем, в 10 из 12 изученных в Европе и Северной Африке у чёрного дрозда Turdus merula (Müller et al., 2013b); 4 — у большой синицы в Дании (Müller et al., 2013a), потом также и в Барселоне (Riyahi et al., 2015, 2017) и др. Поэтому их разумно рассматривать как приспособление к самым общим характеристикам урбосреды, определяющим её экстремальность для «диких» видов птиц и других позвоночных.
Недавно опубликованы крайне подробные данные по генетике урбанизации больших синиц в разных местах европейского континента. С одной стороны, они подтверждают вывод об общих тенденциях микроэволюции при освоении урбосреды, дополняя их массой интересных подробностей:
а) птицы отбираются прежде всего по поведению — как при доместикации, но с иными последствиями, а поведенческие изменения «тянут за собой» остальное (морфофизиологию, генетику и т. д.);
б) разные города заселяются независимо из ближайших участков природных биотопов, без «прыжков» урбанизированных популяций в незанятые ими города,
в) ДНК-филогения показывает, что давно возникшие (и сильно специализированные, прошедшие жёсткий отбор и т.д.) городские популяции разных видов птиц, млекопитающих, ящериц и т.д. ближе каждая к исходной популяции в «своём» ближнем пригороде, а не друг к другу, т. е. при расщеплении популяционной системы вида в связи с урбанизацией все «городские» особи быстро (и однотипно) специализируются к устойчивому существованию в урбосреде, но не обосабливаются так, что их можно выделить в отдельную, например, географическую расу, не говоря уж о виде, а птицы из разных городов не делаются генетически более сходными между собой,
г) общий тренд изменений поведения, физиологии, биоритмов и пр. при урбанизации существует, но по-разному реализуется в разных городах, в том числе разными генами. В частности, у больших синиц это не везде связано со знаменитыми «генами урбанизации», DRD4 и SERT.
P.Salmon et al. (2021) генотипировали 192 больших синиц Parus major из 9 европейских городов и сельских местообитаний той же региона. Отобранные в разных городах гаплотипы совпадают редко, но типичные для городов изменения происходят внутри одних и тех же генов, в основном связанных с нейронной функцией и развитием. К сожалению, в случае приспособления к городской среде короткий эволюционный временной масштаб, зависимость эволюции от местных факторов и полигенная природа многих фенотипических признаков затрудняют обнаружение геномных сигналов адаптации.
Объём данных составлял 10-16 особей в каждой выборке, 517603 SNP. За счёт изучения неравновесия по сцеплению была оценена структурированность европейской популяции, она оказалась низкой. При анализе генетической вариабельности методом главных компонент первые две оси объясняли по 3% изменчивости. В городских популяциях не наблюдается снижения гетерозиготности, т.е. они не проходили сквозь бутылочное горлышко: это независимое подтверждение их формирования направленно «сортировкой» особей по поведенческим потенциям между контрастирующими ландшафтами.
Другое подобное подтверждение — множественные различия «городских» и «лесных» экотипов не объяснимы дивергентным отбором на местах размножения. Данные были получены на синицах, гнездящиеся в дуплянках в г.Монпелье vs в дубовом лесу в ближнем пригороде (Caizergues et al., 2018). Между группировками выявлены четыре устойчивых различия в морфологии (в городе короче цевка, меньше длина крыла и хвоста, ниже масса тела) и два — в жизненной стратегии (более раннее размножение, меньший размер кладки). Давления отбора между дивергентными биотопами были дивергентны только для массы тела, по иным признакам каждой группировке присущ свой набор селективных давлений, не связанных между собой (Caizergues et al., 2018). .
Уровни миграции за пределами центральной части выборки, оцененные Salmon et al. (2021), обычно низкие, в том числе между близкими городами. Некоторые городские популяции кластеризуются вместе со своими «лесными» парами. Тем не менее, анализ предполагает определённый уровень миграций, в том числе дальних, например из Лиссабона в Глазго. Генетическая дифференциация населения (FST) внутри пар популяций («городское» vs ближайшее «лесное» население), и между населением разных городов в среднем выше, чем между «лесным» населением разных районов Европы. Видим, что поток генов обычно сильнее между популяциями исходных биотопов, чем между городскими (в 9 парах подобных сравнений); главное, что он резко снижается при их отщеплении от «лесных».
Предыдущие результаты на другом виде воробьиных Европы, черном дрозде, показали, что разные города колонизируются им независимо из ближайших лесов. В данной работе обнаружение независимых кластеров для некоторых городских популяций также указывает на аналогичный сценарий, то есть на независимую и повторяющуюся колонизацию городов преимущественно смешанными популяциями в исходных биотопах. Однако общая слабая генетическая дивергенция и обнаружение значительных миграционных событий среди отдаленных популяций предполагает возможную роль потока генов в облегчении городской адаптации через распространение адаптивных аллелей (Salmon et al., 2021).
2758 SNP, связанных с урбанизацией согласно регрессионной модели LFMM (latent-factor mixed models), были распределены по всему геному и не группировались в определенных регионах. Метод главных компонент чётко разделяет «лесные» и «городские» популяции (доля объясненной дисперсии PVE — PC1 = 1,98%). Результаты, полученные байесовским подходом (BayPass), выявили лишь 70 SNP, сильно связанных с урбанизацией, из них 34 совпали с выявленными LFMM (много больше случайного). Среди этих 34 SNP частота одного и того же аллеля увеличена в семи или более городских популяциях по сравнению с парными сельскими. Авторы показали, что существует локальная адаптация к городской среде обитания у больших синиц, и что это происходило по всей Европе за счет сдвигов в частоте аллелей одних и тех же локусов (Salmon et al., 2021).
Это могло произойти, поскольку предположительно адаптивные аллели были общими для значительной части европейского ареала вида, или из-за обмена адаптивными аллелями через поток генов между городскими популяциями. Тем не менее, в соответствии с полигенной основой адаптации посредством тонких сдвигов частоты аллелей в нескольких локусах, обнаруженные различия между городом и лесами вокруг в целом невелики. Только несколько SNP показывали относительно сильные сдвиги частоты аллелей (>0.5). Далее авторы искали участки генома с большим числом отличий между «городской» и «лесной» популяцией. Их было довольно много, хотя они слабо коррелировали между разными городами (были общими максимум для пяти пар популяций). Три наиболее значимых общих участков такого рода были сгруппированы в области размером 300 тыс. п.н. на хромосоме 11 (16,05–16,35 МБ) и отмечены в Гётеборге, Мюнхене и на Пиренеях (Барселона, Мадрид и Лиссабон).
В целом число совпадений не связано с географической близостью популяций. Например, максимум общих участков (53 и 47 соответственно) фиксируется в Гётеборге и Париже, а также в Лиссабоне и Глазго. Таким образом, общие адаптивные генетические вариации через интрогрессию и поток генов между соседними городскими популяциями вряд ли вносили существенный вклад в адаптацию (Salmon et al., 2021).
Расхождения результатов, полученных разными методами обнаружения «подписей» (англ. signature) естественного отбора5, говорят о том, что большинство селективных изменений недавние и незавершённые. Авторы также проверили, что последние действительно вызваны отбором, а не генетическим дрейфом или попутным транспортом в регионах с низкой рекомбинацией – в том числе потому, что некоторые участки, для которых наблюдался отбор, располагались в областях с высоким уровнем рекомбинации. Для влияния генетического дрейфа генетическая дифференциация популяций слишком мала; кроме того, помимо Глазго и Лиссабона (краевые популяции) практически не обнаруживалось дефицита гетерозигот.
Дальше исследователи сосредоточились на генах, связанных с признаками отбора более чем у половины всех «городских» популяций (≥5 городских популяций): они, вероятно, важнее всего для адаптации к урбосреде. Они обнаружили 42 гена, ассоциированных с недавним или продолжающимся отбором, и 15 других, связанных с более ранними и завершёнными событиями отбора, но не отметили роста числа общих генов под отбором в зависимости от географической или генетической дистанции между птицами, подтвердив независимость многих событий освоения разных городов. Кроме того, 12 из 57 общих генов при отборе также были связаны с урбанизацией в анализе LFMM. Интересно, что два из них, GMDS и SLC6A15 по-разному экспрессируются в крови и/или печени между городскими и сельскими птицами (Salmon et al., 2021).
Несмотря на наличие «подписей» отбора, перекрывающих один и тот же ген в пяти или более городских популяциях (например, гены GMDS, HTR7 или CDH18), их точное расположение не соответствовало друг другу. Более того, в некоторых случаях самые сильные «подписи» воздействия отбора находятся не внутри генов, а вверх или вниз по течению6 от них, и не имеют согласованного паттерна среди популяций. Например, признаки отбора вокруг HTR7 присутствовали вниз по течению в Глазго и Мюнхене, но вверх – в Мальмё, Париже, Мадриде и Лиссабоне. Это может свидетельствовать о том, что с адаптацией к городу связаны изменения именно в регуляторной, а не белок-кодирующей части генома, причём их природа различна в разных популяциях.
Многие из генов, связанных с урбанизацией, ранее были связаны с поведенческой дивергенцией, а также с когнитивными функциями и обучением. В частности, ген HTR7 (хромосома 7) кодирует рецептор нейромедиатора серотонина, путь, который последовательно идентифицируется как цель изменений, связанных с урбанизацией. Действительно, два предыдущих исследования (на европейских черных дроздах с использованием подхода кандидатных генов и на кроличьих сычах с использованием полногеномного ресеквенирования) также описали изменения в этом пути как основное различие между «городскими» и сельскими популяциями. Оба вида также демонстрируют постоянные различия между данными популяциями в двух формах поведения, связанных с серотонином: подавление активности «оси стресса» и её продление активности вечером и ночью при искусственном освещении. HRT7 также напрямую связан с различиями в поведении мигрирующей и жилой радужной форели Onchorynchus mykiss.
Ген CDH18 (хромосома 2) — часть суперсемейства мембранных белков, участвующих в синаптической адгезии, выявлен как ген-кандидат в фонологических изменениях у людей. Предполагается, что гены PTPRD и VPS13A (оба в Z-хромосоме) участвуют в развитии гиппокампа и ранее связывались с навигацией птиц, полётной активностью и миграционным поведением. Более того, два других гена, находящихся в процессе отбора и связанные с урбанизацией, DLG2 и NRXN3 в хромосомах 1 и 5, участвуют в регуляции когнитивных процессов и поведенческих расстройств: ранее их связали с отличиями «городских» кроличьих сычей (Salmon et al., 2021).
Эти результаты дополнительно подтверждены анализом 2758 связанных с урбанизацией SNP (анализ LFMM), связанных с 984 генами (1501 SNP в генных регионах), с точки зрения принадлежности последних к определённым функциональным терминам Gene Ontology (задают категории белков, исходя из их функции в клетке, ткани или организме). Те из них, что перепредставлены при урбанизации, связаны в основном с функционированием и развитием нейронов, клеточной адгезией и сенсорным восприятием, и сгруппированы в две взаимодействующие сети, одна из которых связана с сенсорным распознаванием, другая — с развитием нервной системы и клеточной адгезией. Эти результаты подтверждают предыдущую идею о когнитивных и поведенческих изменениях как ключевых ответах на урбанизацию. Цитируемое исследование предполагает наличие сильного генетического компонента этих предположительно городских адаптивных фенотипов у больших синиц по всей Европе (Salmon et al., 2021).
Другая работа по большой синице (Caizergues et al., 2022) выполнена на трёх парах «городская популяция — популяция из соседнего лесного массива» Барселоны, Монпелье и Варшавы. Из каждой брали по 10 особей, в образцах крови анализировали как различия по частоте SNP, так и DMR (дифференциально метилированные регионы генома). Обнаружены признаки слабого отбора, во всех случаях «разводящего» городские и лесные популяции. DMR преимущественно попадают на сайты транскрипции и промоторные области, что также свидетельствует об их значимости. Как SNP, различающиеся по частоте в городских и лесных популяциях, DMR часто попадали в локусы, связанные с нервной системой, иммунитетом, поведенческими, гормональными и стрессовыми реакциями. Однако параллелизм по конкретным локусам для трёх пар популяций весьма незначителен.
Из генов, меняющихся в связи с приспособлением к урбосреде, выявлены NR4A2, значимый при распознавании новых объектов и памяти у мышей; DCX, связанный с пластичностью нейронов. В эксперименте с воронами показано, что искусственный ночной свет вызывает его гиперэкспрессию, с изменением поведения и появлением депрессивноподобного поведения разной выраженности. Наконец, это ген CHRNA1, ассоциированный с агрессией у кур. Найдены четыре гена (POMC, ADAMTS3, PAPD4 и GCC1), связанные с DMR, которые ранее у больших синиц описывались как серьезно меняющие уровень метилирования под действием загрязнений (их точные функции пока неизвестны). Однако не выявлено DMR, связанных с известными «генами урбанизации» SERT и DRD4, хотя обнаружено сильное изменение метилирования DRD3 (связан с агрессивным поведением кур). Следует учитывать, что судя по предыдущим исследованиям, метилирование сильно отличается в зависимости от выбранной ткани и подвержено сезонным изменениям (Caizergues et al., 2022).
Примечания
1 Особи, совершающие инвазию в нетипичный ландшафт (за границей видового ареала и/или в экстремальные среды обитания внутри него — обычно в изменённых человеком ландшафтах, особенно урбо-) почти всегда гибнут полностью. Скажем, при повторяющихся налётах больших пёстрых дятлов Dendrocopos major в тундру и другие безлесные ландшафты (инвазий) их участники массово гибнут от голода или (как на Ямале) истребляются сапсанами Falco peregrinus (Галушин, 1962, цит.по: Бутьев, Фридман, 2005). Поэтому после интенсивных инвазий, вовлекающих взрослых птиц, наравне с молодыми, у соответствующих видов возникает депрессия численности.
Элиминация дятлов в тундре сравнима с массовой гибелью мелких птиц, привлекаемых светом больших городов, особенно во время миграций. “Притяжение” к городам пропорционально освещённости ночного неба (Horton et al., 2023): пересекающие их особи бьются сотнями тысяч о стеклянные поверхности и/или освещённые стены домов (Loss et al., 2014, русский пересказ см. «Освещение города и гибель птиц»). Проведение световых шоу, с резкими движениями этой подсветки — дополнительный фактор гибели, увеличивающий её значительно больше базового уровня, заданного общей освещённостью. Ещё важнее, что уже как минимум 50 лет «диким» видам во время перемещений никак «не уклониться» от пересечения урболандшафта: с 1960-х гг. города из внемасштабных образований стали пятнами, выражаемыми в масштабе карты и, главное, большими по размеру, чем лесные, луговые или болотные массивы того же региона (Фридман, Суслов, 2022).
Однако при успешной урбанизации тех же видов, осваивающих города, которые раньше лишь пересекали при перелётах, не происходит сколько-нибудь значимого подъёма гибели даже на первых “шагах” освоения. По его завершении этих же (как и близких видов) не несут заметных потерь от этого и других факторов риска, хотя постоянно живут среди зданий, стеклянных поверхностей, проводов и пр. источников риска, а не просто за короткое время пересекают их. Больше того, их СОПЖ увеличена сравнительно с исходными («лесными» или «сельскими») популяциями — настолько, что её удлинение «перевешивает» сокращение ежегодного успеха размножения составляющих её птиц, так что репродуктивный выход «городских» популяций выше, чем исходных (Фридман, Суслов, 2022). Это один из симптомов успешного приспособления к урбосреде.
2Мажорирование — приём рассуждения, состоящий в заведомом усилении тезиса, защищаемого или опровергаемого говорящим, так, чтобы затрудить доказательство первого или второго. Но если последнее всё же получится, это придаст максимальную убедительность эмпирическим данным, использованным для локазательства, и данному выводу из них в сравнении с другими возможными, но оставшимися не обсуждёнными.
3См.пример «самоспасения» голубя-клинтуха Columba oenas, начавшего гнездиться в бетонных столбах ЛЭП среди агроландшафта вместо прежде лесного образа жизни, с гнездованием в дуплах, обычно выдолбленных желной Dryocopus martius.
4CPG-острова — участки генома с повышенной частотой встречаемости динуклеотидов СG, выделенные по некоторому верхнему проценту отсечения. Цитозины в них часто метилируются с образованием 5-метилцитозина, что на порядок увеличивает шансы мутаций, превращающих его в тимин. Поэтому геном вообще-то парами CG обеднён; там, где они концентрируются — в CpG-островах — скорей всего, действовал отбор, обеспечивший их консервативность. Соответственно, «острова» обычно указывают на начало транскрипции, выступая альтернативой ТАТА-боксу, и метилируются обычно в активно экспрессирующихся генах. Метилирование СG-пар недалеко от «острова» влияет на уровень экспрессии гена.
5Как это делают биоинформатики, см. «Быстрая эволюция к безбивневости у африканских слонов вследствие браконьерства».
6Направление задано «правилами чтения» генетического текста, от 3` к 5` концу нити ДНК.
Поведенческие изменения у «городских» чечевичников «тянут» за собой морфологию
Красивый пример амальгамы прямого приспособления и наследуемых изменений, чей общий фенотипический результат закрепляется отбором (включая устойчивую реализацию в онтогенезе) дают «городские» популяции мексиканского чечевичника Haemorhous mexicanus. У самых разных видов певчих птиц урбанизация сопрягается с ростом частоты вокализаций, составляющих песню, чтобы их «вывести» из полосы техногенного шума, мешающего потенциальным партнёрам распознавать её видоспецифическую структуру (Gil, Brumm, 2014; Halfwerk, Slabbekoorn, 20141). У так называемых «толстоклювых» воробьиных эта частота определяется формой клювного конуса: на примере мексиканского чечевичника исследовали, меняет ли то и другое урбанизация, анализируемая на градиенте переходов от «сельских» местообитаний к «городским» (Giraudeau et al., 2014).
Оказывается, да, у птиц на более урбанизированных территориях клюв делался тоньше и длинней, что повышало частотный диапазон пения и максимальную частоту складывающих его звуков. Как в примерах описанных выше, здесь поведенческие изменения «тянут за собой» морфологические, причём пошедшие через 10 и более лет после формирования городской популяции, когда она уже показала свою жизнеспособность, растёт численно и изменила свою «естественную историю» (Giraudeau et al., 2014).
Самки чечевичников предпочитают самцов, поющих больше песен, и с более длинными песнями. Поэтому важно, чтобы их видоспецифическое звучание не маскировал городской шум (низкочастотный): здесь могут помочь изменения размера и формы клюва, включая угол его раскрытия. Голосовой тракт, трахея, гортань и клюв при пении работают как резонансная камера, а тонкие изменения размера/формы клюва и морфологии голосового тракта сильно влияют на издаваемые звуки. Обычно более длинный, меньше открывающийся, но более широкий клюв обеспечивает значимо меньшие минимум и максимум частоты пения, а также полосу пропускания.
Авторы изучили связь формы клюва и частотной структуры пения мексиканских чечевичников в зависимости от степени урбанизации территорий, лежащих вокруг г. Финикс (Аризона, США); их ряд образует городской градиент.
Степень урбанизации каждой из них оценивали по плотности населения и семи измерениям, касающимся разных паттернов землепользования в 1 км вокруг места поимки птиц (таблица 1). Для численной характеристики степени урбанизации названные 8 метрик анализировали методом главных компонент (PC). Были выделены три координатных оси, PC1-3, вместе объясняющие >84% изменчивости ландшафта, заселенного чеченвичниками: PC1 — 47% дисперсии, PC2 и PC3 — по 24% и 13%. PC1 сильно и негативно коррелировал с % земель, занятых нативным ландшафтом пустыни (вклад этой оси >94%). PC2 сильно и коррелировал с % земель, занятых культивируемой растительностью (вклад >81%), PC3 с % земель, занятых местной растительностью (вклад >74%) (Giraudeau et al., 2014).
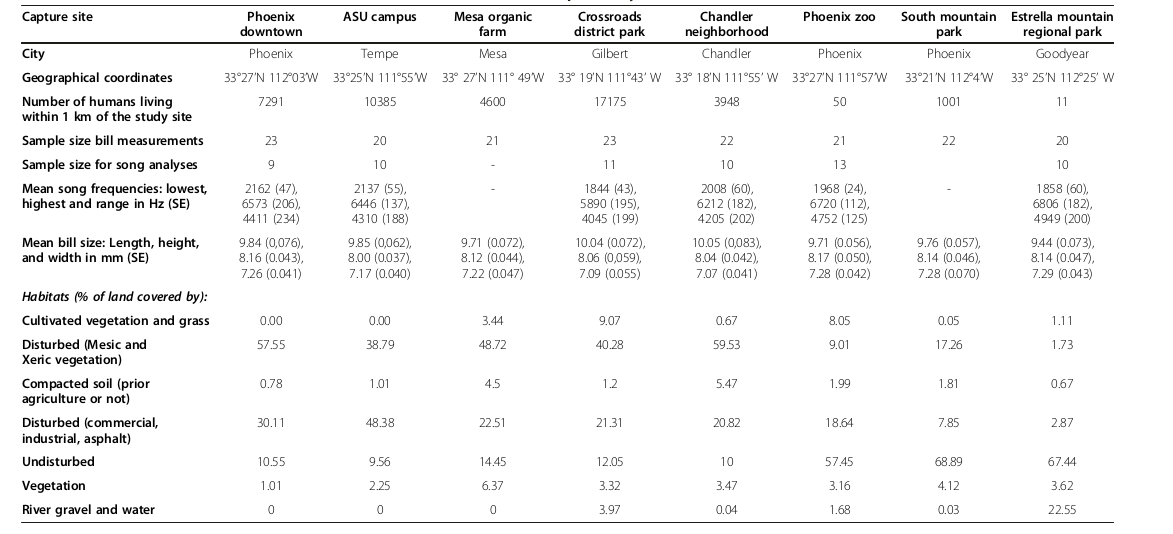 Таблица 1. Изменения формы клюва и песенных характеристик вдоль градиента урбанизации близ г.Финикса: данные, использованные для анализа.
Таблица 1. Изменения формы клюва и песенных характеристик вдоль градиента урбанизации близ г.Финикса: данные, использованные для анализа.
Обозначения. Сверху вниз: населённый пункт, географические координаты, плотность населения в радиусе 1 км, объём выборки для измерений кл.юва, объём выборки для анализ структуры песен, средние характеристики частотной структуры видовой песни: максимум, минимум, диапазон между ними (кГц), структура ландшафта в радиусе 1 км вокруг, % площади, покрытой: возделываемой растительностью, с/хугодья; антропогенно нарушенных (мезо- и ксерофильная растительность); уплотнённой почвой (бывшей ранее в с/хиспользовании или нет); застроенные территории (коммерческая и индустриальная застройка), запечатанные поверхности почвы (асфальт); незастроенные территории; общее покрытие растительности; водное зеркало и песчаные отмели.
Длина цевки и масса тела вьюрков не коррелировали с морфометрией клюва (все P > 0,39). Однако длина цевки (rho = 0,76, P = 0,03) коррелировала с PC2: она удлинена у птиц, отловленных на территориях с большими площадями возделываемой растительности. Форма клюва зависит от степени урбанизации: длина положительно связана с РС1, ширина и высота отрицательно (rho = 0,76, P = 0,03; −0,79, P = 0,02; −0,52, P = 0,18). Чем сильнее пустыня была трансформирована в урбо- или с/хландшафт (т. е. вдоль городского градиента), тем сильней увеличивалась длина клюва чечевичников, а ширина и высота уменьшались (рис.1). Ни один из параметров формы клюва не коррелирует с координатами PC2 и PC3 (все P > 0,49).
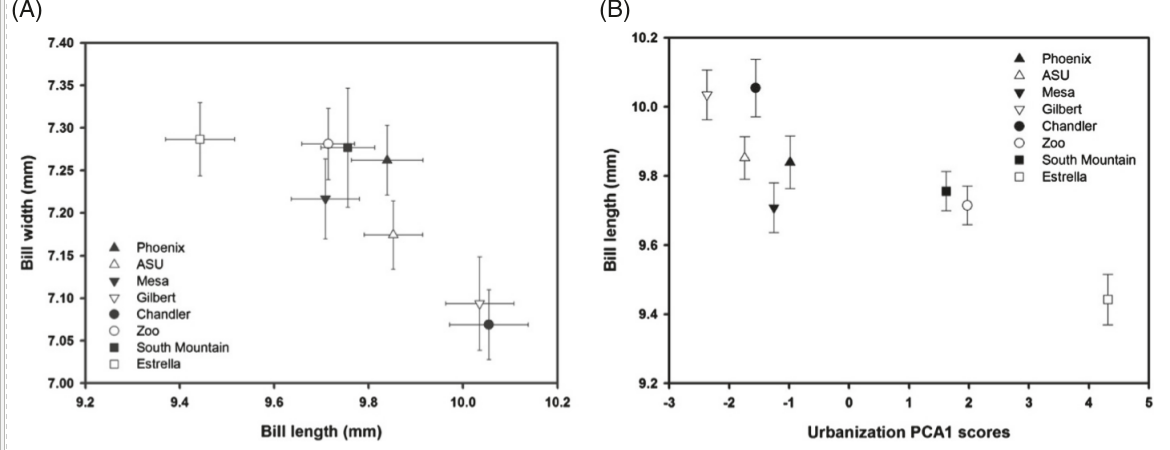 Рисунок 1. Изменения клювного конуса мексиканских чечевичников в зависимости от урбанизованности территории. А. Связи параметров клювного конуса (длина — абсцисса, ширина — ордината, всё в мм) на 8 изученных территориях вокруг Финикса (значки слева внизу). В. Зависимость длины клюва (ордината, мм) от РС1 для тех же 8 участков. Здесь и дальше источник Giraudeau et al., 2014, fig.1-4.
Рисунок 1. Изменения клювного конуса мексиканских чечевичников в зависимости от урбанизованности территории. А. Связи параметров клювного конуса (длина — абсцисса, ширина — ордината, всё в мм) на 8 изученных территориях вокруг Финикса (значки слева внизу). В. Зависимость длины клюва (ордината, мм) от РС1 для тех же 8 участков. Здесь и дальше источник Giraudeau et al., 2014, fig.1-4.
В ряду названных 8 территорий средние ширина и длина клюва связаны отрицательно (rho = -0,86, P = 0,006) при положительной связи ширины с высотой (rho = 0,76, P = 0,03). Последняя не коррелирует с длиной клюва на уровне всей популяции (rho = -0,55, P = 0,16).
Урбанизация и структура песни вьюрков сопряжённо меняются вдоль городского градиента
В ряду из 8 территорий средние значения максимальных частот фонового шума отрицательно коррелировали с PC2 (rho = -0,83, P = 0,04), но не с PC1 (rho = 0,03, P = 0,96) или PC3 (rho = 0,60, P = 0,21). Иными словами, сильнее зашумлены территории, меньше покрытые растительностью. В том же ряду самые низкие частоты звуков, образующих видовую песню H.mexicanus, значимо положительно коррелировали с максимумом частоты фонового шума (rho = 0,94, P = 0,005; рис.2); никакие другие характеристики песен не с зашумленностью не связаны (все P>0,6). Минимум частоты пения также отрицательно коррелировал с PC2 (rho = -0,94, P = 0,005), тогда как максимум (rho = -0,83, P = 0,04) и ширина диапазона между ними (rho = -0,84, P = 0,04) коррелируют с РС1 положительно. Вьюрки пели на более высоких частотах в местах, менее покрытых растительностью, и снижали максимум частоты/сужали частотный диапазон на более трансформированных участках.
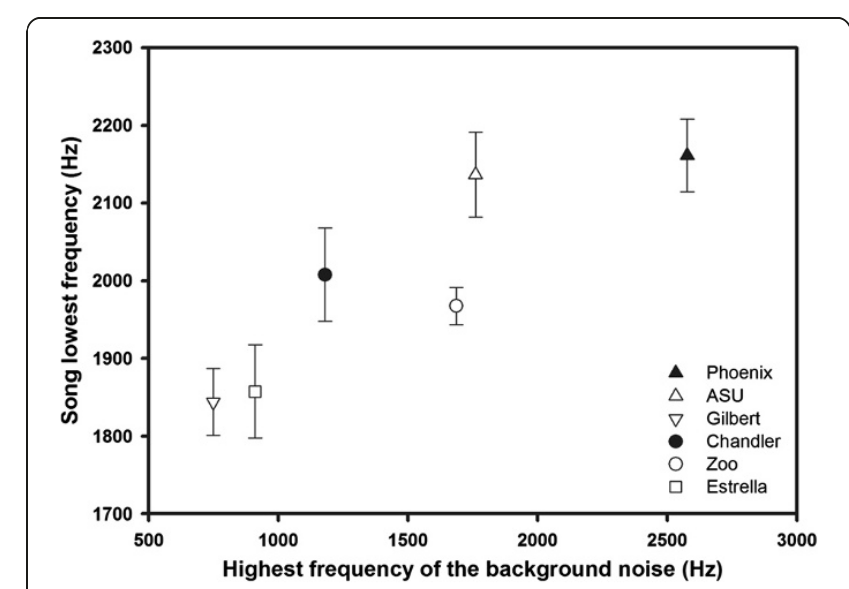 Рисунок 2. Положительная связь минимальной частоты пения и максимальной частоты фонового шума в ряду 6 территорий, для которых можно было собрать данные о структуре песни. Показаны средние ±SE.
Рисунок 2. Положительная связь минимальной частоты пения и максимальной частоты фонового шума в ряду 6 территорий, для которых можно было собрать данные о структуре песни. Показаны средние ±SE.
Сопряжённое варьирование формы клюва и характеристик песни вдоль городского градиента
Различия средней длины и ширины (но не высоты, Р>0,05) клюва вьюрков в ряду территорий значимо связаны с характеристиками их пения. У птиц с более длинными и узкими клювами (это, как помним, связано с трансформированностью ландшафта) ниже максимум песенных частот (rho = -0,94, P = 0,005; рис.3) и уже частотный диапазоном звуков, представленных в песне (rho = -0,94, P = 0,005; рис.4). Самые низкие частоты пения птиц не были связаны с морфологией клюва (все P>0,8). Другими словами, изменения формы клюва, связанные с «городской жизнью», коррелировали с изменением максимума частоты пения и диапазона частот (Girardeau et al., 2014).
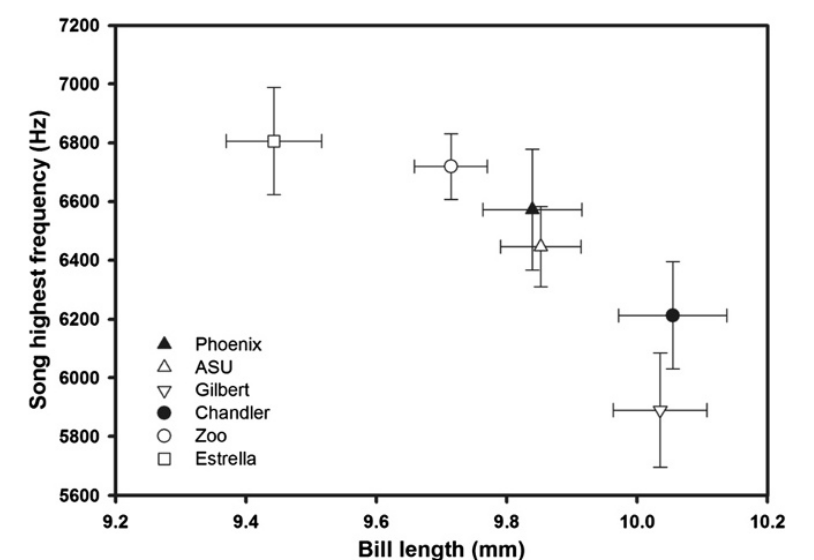 Рисунок 3. Зависимость между максимальными частотами пения (кГц, ордината) и и средней длиной клюва (мм, абсцисса) в ряду 6 территорий (значки слева внизу), для которых получены данные о том и другом признаке (±SE).
Рисунок 3. Зависимость между максимальными частотами пения (кГц, ордината) и и средней длиной клюва (мм, абсцисса) в ряду 6 территорий (значки слева внизу), для которых получены данные о том и другом признаке (±SE).
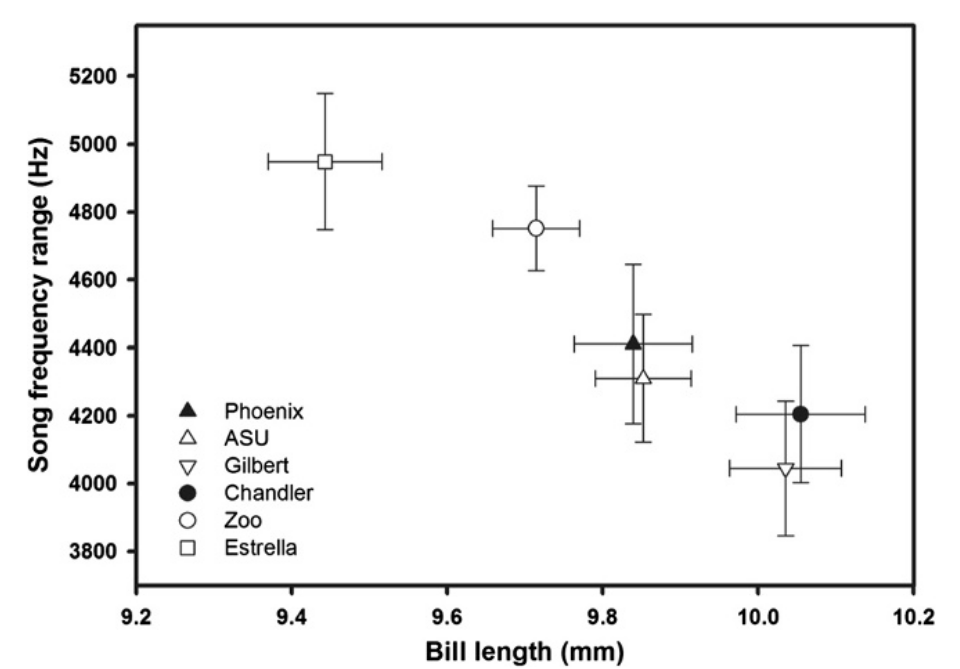 Рисунок 4. Зависимость между шириной частотного диапазона песенных звуков (кГц, ордината) и средней длиной клюва (мм, абсцисса) в ряду 6 территорий (значки слева внизу), для которых получены данные о том и другом признаке (±SE).
Рисунок 4. Зависимость между шириной частотного диапазона песенных звуков (кГц, ордината) и средней длиной клюва (мм, абсцисса) в ряду 6 территорий (значки слева внизу), для которых получены данные о том и другом признаке (±SE).
Обсуждение
Рост степени урбанизации территорий меняет форму клюва чечевичников, он делается длиннее и уже. Обычно различия в форме клюва городских и “сельских” птиц объясняют шаблонно, разным составом кормов. Мол, в городах на кормушках они едят подсолнечник и арахис (более крупные семена с более твёрдой оболочкой), вне города — семена кактусов или сорняков, мелкие, с более тонкой кожурой. Чтобы с первыми справиться, клюв должен быть крупнее, с более сильным укусом. Однако, это не объясняет, почему в Финиксе клювы «городских» чечевичников тоньше и длиннее, чем у «сельских». Самое правдоподобное объяснение — удлинение клюва даёт преимущество в обработке крупных семян на кормушках, а для обработки мелких семян в сельской местности больше годится клюв покороче. Действительно, воробьиные с самыми короткими клювами быстрее всего шелушат самые мелкие семена, более крупноклювые виды быстрее чистят самые крупные (Soobramoney, Perrin, 2007, цит.по: Girardeau et al., 2014).
Точней, изменëнная форма клюва рождается изменением процессов пролиферации и миграции тканей будущего клюва в онтогенезе. Он же в значительной степени регулируется экспрессией костных морфогенетических белков (BMP) на ранних стадиях развития. Если в более урбанизированных районах молодые вьюрки лущат семена с другим уровнем механического воздействия на клюв, то это влияет на выработку BMP, через них — на рост клюва на протяжении целых 2,5 месяцев после вылупления. Недавно Badyaev et al. (2008, цит. по: Girardeau et al., 2014) показали, что разные уровни экспрессии BMP чечевичников в городе и в пустыне могут установиться еще до вылупления. Возможно, [стабилизирующий] отбор рационализировал онтогенез “городских” вьюрков настолько, что поднял уровни экспрессии белков BMP ещё у эмбрионов в яйце, обеспечив формирование у них более длинных и тонких клювов (Girardeau et al., 2014).
Закономерные изменения формы клювного конуса вдоль городского градиента также связаны с изменением структуры видовой песни, прежде всего её частотных характеристик. Более длинный и узкий клюв птиц с более антропогенно трансформированных территорий жёстко связан с уменьшением максимальной частоты пения. Это хорошо согласуется с данными о снижении частоты звучания песен, создаваемых в более длинном голосовом тракте, в том числе за счёт удлинения клюва. Однако минимальная частота пения не снижается, вопреки удлинению клюва с ростом урбанизованности территории, т. е. шире частотный диапазон, в котором звучат песни «более городских» особей. Больше того, максимум уровня фонового шума сильно и положительно коррелирует с минимальной частотой пения чечевичников, что обеспечивает эффективность коммуникации в городе, хотя, будучи более длинноклювыии, в отсутствие необходимости экранировать шум эти птицы могли бы петь сильно ниже (Girardeau et al., 2014).
Видим, что в освоении урбосреды чечевичниками реализон адаптивный компромисс, позволяющий изменяться под одновременным действием двух взаимно противоречивых селективных давлений. Корм из более крупных/жестких семян делает клюв длиннее и тоньше, но не всеми последствиями этого для частотного диапазона пения можно воспользоваться вследствие роста зашумленности в том же ряду территорий. В результате реализованный диапазон у “городских” птиц на 20% уже, чем у пустынных. В эту же сторону “работает” индивидуальная пластичность “выдачи” песенной продукции: столкнувшись с дополнительно созданным шумом или его уменьшением, одни и те же самцы адекватно меняли минимальную частоту пения (Bermúdez-Cuamatzin et al., 2011, цит.по: Girardeau et al., 2014).
Увы, авторам неизвестны каких-либо исследования, показывающие пластичность исполнения на максимальных частотах в ответ на городской шум. В будущих экспериментах следует изучить роль генов, индивидуального развития, обучения и пластичности голосовых путей в формировании разнообразных вокальных характеристик в антропогенной среде.
Далее авторы задаются вопросом: каковы могут быть экологические и эволюционные последствия сопряжённых изменений песни и формы клюва по ходу урбанизации? Особенности песен являются ключевыми показателями качества самцов у многих видов птиц [47]. У домового вьюрка самки продемонстрировали значительные предпочтения в выборе партнера, основанные на характеристиках песен [34]. Следовательно, уменьшение частотного диапазона сигнала в ответ на активность человека может иметь серьезные репродуктивные последствия для мужчин. С другой стороны, гибкость в выборе самок может позволить приемникам использовать альтернативные вокальные компоненты, которые более достоверно отражают мужские качества в новой среде [48],[49]. В соответствии с этой гипотезой, Halfwerk и соавт. (2011, [49]) недавно экспериментально продемонстрировали преимущество самцов больших синиц (Parus major) в передаче сигналов при высокочастотных песнях в шумных условиях, в то время как низкочастотные песни, вероятно, предпочтительнее в естественной, менее шумной среде.
Примечания
Рекомендуем прочесть
 Городские ящерицы любят погорячее, и их гены объяснят, почему
Городские ящерицы любят погорячее, и их гены объяснят, почему «Культурная передача» навыка открывания крышечек у лазоревок
«Культурная передача» навыка открывания крышечек у лазоревок Регенеративная урбанизация: Экополис
Регенеративная урбанизация: Экополис Урбанизация и антропогенез: неожиданные параллели
Урбанизация и антропогенез: неожиданные параллели Глобальное потепление загоняет соловьёв в ловушку
Глобальное потепление загоняет соловьёв в ловушку Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки
Человеческая деятельность формирует зимнюю экологию перелётной птицы, славки-черноголовки Как соотносятся прямое приспособление и эволюционные изменения?
Как соотносятся прямое приспособление и эволюционные изменения? Ещё про гены и поведение
Ещё про гены и поведение
