
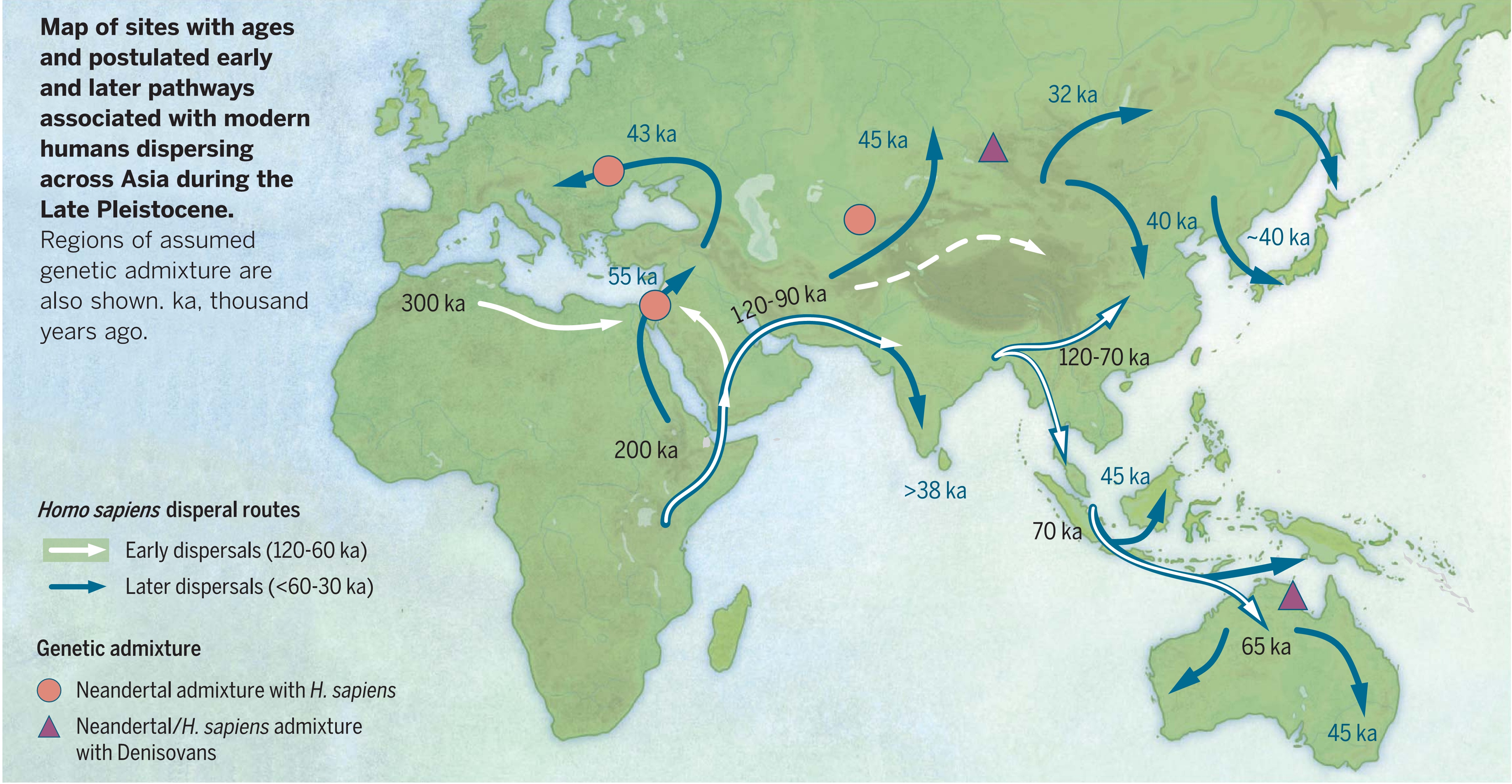
Пути миграций Homo sapiens: белыми стрелками обозначены ранние миграции (120-60 тыс. лет назад), синими стрелками поздние миграции (60-30 тыс. лет назад). Розовые кружки показывают места метисации сапиенсов с неандертальцами, треугольники — места метисации с денисовцами. Источник Генофонд.рф
В их основе — инвазия в экстремальную среду обитания
Содержание
- 1 В их основе — инвазия в экстремальную среду обитания
- 2 Урбанизация и антропогенез: параллель генетических изменений
- 3 Урбанизация меняет индивидуальность: роль DRD4 и SERT
- 4 Полиморфизм DRD4 влияет на восприятие новизны у птиц и млекопитающих
- 5 Антропогенез как всё лучшее использование новизны во всё более экстремальной среде обитания
- 6 Заключение
Обсуждая урбанизацию «диких» видов птиц и млекопитающих, учёные рассматривают два механизма, отвечающие на вопрос «что именно изменяется, для чего и зачем?». Первый ответ, сегодня очень популярный на Западе — это своего рода одомашнивание, сходные изменения описали при одомашнивании лисиц, норок и пр. Д.К.Беляев, Л.Н.Трут и другие в Новосибирске (одно из немногих достижений советской науки, не замолчанное и не присвоенное там, а, наоборот, уважительно цитируемое). Мол, «городские» чёрные дрозды, большие синицы, вяхири и т.д. становятся «добрей и доверчивей» к людям, как лисицы в беляевском домике, у первых меняется то же и так же, что у вторых (рис.1).
Второй ответ предлагается нами с В.В.Сусловым из ИциГ СО РАН: при освоении городов «дикие» виды птиц и млекопитающих совершают инвазию в экстремальную среду обитания, и приспосабливаются к её наиболее общим параметрам (нестабильности, изменчивости и гетерогенности немыслимой и нетерпимой в их прежних местообитаниях). Потому у них изменяются физиология и генетика (отбор по «генам урбанизации», они же «гены индивидуальности» DRD4 и SERT) в сторону позитивного отношения к новизне, лучшей дифференцировке происходящего вокруг в условиях беспокойства (в том числе его отделения от опасности), большей точности действий в этих условиях. Это очень похоже на антропогенез: выходя из тропического леса в саванну, позже распространяясь из Африки (а также из «колыбели» Восточной и Южной Африки в Западную и Центральную) человек, подобно «городским» птицам и млекопитающим, ввергал себя в экстремальную среду — новую, исторически нетипичную, взаимодействие с которой влечёт за собой неизвестные риски, но и содержит возможности, оправдывающие её освоение.


Б.

В.
Рисунок 1. Разные аспекты доместикационного эксперимента. А. Д.К. Беляев с ручными лисами у домика. Б. Крайнее выражение страха перед человеком у «дикой» лисицы. Видно отсутствие выбора: животное всё бросает, лишь бы уйти подальше, но боится повернуться спиной к человеку, чтобы залезть выше на сетку. В. Доместицированные лисицы сохраняют это отсутствие выбора: всё бросают, только чтобы бежать к человеку, а не от него.
Источник. Колчанов Н.А. Регуляторные генетические системы: экспериментально-компьютерный анализ и моделирование.
Быстрое развитие мозга и интеллекта здесь было возможно лишь вместе с принятием новизны (воспринимаемой как потенциальная опасность и/или помеха, а то и являющейся ею), готовностью действовать ей вопреки, тогда как раньше те же воздействия «автоматически» вели к отступлению или бегству. Освоение новой, исторически нетипичной среды требовало способности устойчиво действовать в ней, действовать» «вопреки» факторам экстремальности (как конкретным воздействия вроде шума транспорта, людей и собак, или общим отличиям от исторически типичной в виде аномально высокой изменчивости, нестабильности, гетерогенности и «кружева» биотопов, и условий существования в них), в том числе действовать несмотря на временное отсутствие результата, способность достигать цели долговременных программ поведения несмотря на ухудшение ситуации поначалу и т. д. трудности, разрешаемые также и «городскими» популяциями птиц и млекопитающих.
Наконец, демография антропогенеза и урбанизации «диких» видов сходна тем, что рост численности в обоих случаях идёт за счёт удлинения среднеожидаемой продолжительности жизни популяции при снижении размножаемости индивидов, только у человека этот эффект выражен всё сильней и сильней на протяжении его истории, а в последние 3-5000 лет и вовсе довлеет, у «городских» птиц и млекопитающих он несравнимо слабей, хоть вполне явственен. И наоборот: адаптация к перманентной нестабильности, гетерогенности, изменчивости новой среды обитания у «городских» птиц, млекопитающих, ящериц идёт тем же способом, если брать лишь «биологию» — изменения на уровне генов, физиологии, жизненных стратегий индивидов. В антропогенезе и при урбанизации «диких видов» это происходит отбором определённых вариантов «генов индивидуальности» DRD4 и SERT (ну и выработкой «орудий социального труда», заражающих позитивом при интеллектуальных усилиях и поднимающих мотивацию действовать несмотря на трудности, к этому «городские» птицы уже неспособны).
Книга «Города как «арены» микроэволюционных процессов» написана в опровержение первой концепции и в доказательство второй. Анализируя данные об экологии и поведении «городских» популяций множества видов птиц, рептилий, млекопитающих, мы показываем, что гипотезу «урбанизации как одомашнивания» они неизменно опровергают, а «урбанизации как инвазии в экстремальную среду», сходную с антропогенезом» — поддерживают и развивают. Поэтому её материал, идеи и выводы книги могут сказать что-то новое и интересное не только про освоение городов «дикими» видами позвоночных, и в таких, кажущихся совершенно несвязанными, области, как антропогенез и «урбанизация растений1», обсуждавшаяся при представлении книги на Ломоносовских чтениях в МГУ.
Урбанизация и антропогенез: параллель генетических изменений
Урбанизация «диких» видов птиц и млекопитающих, с одной стороны, антропогенез — с другой сходны одинаковым направлением отбора полиморфных генов DRD4 и SERT2 (кандидатные «гены урбанизации», они же «гены индивидуальности» у птиц). Мы обозреваем исследования связи генных вариантов DRD4 и SERT с характеристиками поведения и индивидуальности у «городских» птиц и млекопитающих, разных видов приматов, людей с разным образом жизни, в контексте решения разных проблем и болезней, домашних животных, «руководимых» людьми, для определения селективной ценности и реконструкции приспособительного характера соответствующих изменений.
Предполагается, что они прольют свет на особенности антропогенеза, которые остаются невыясненными при обычном генеалогическом подходе, сближающем линию «к человеку» с двумя современными видами шимпанзе, оставляя необъяснёнными системные сходства человека разумного, с одной стороны, с таким «бродяжничающим» и пластичным антропоидом, как орангутан, с другой — с домашними животными. Правда, весьма популярная ныне концепция «самодоместикации человека» не просто ставит последние во главу угла, но доказывает, что системные сходства между Homo sapiens и одомашненными им видами дают полное и исчерпывающее объяснение антропогенеза, как в отношении движущих факторов, так и последовательных изменений морфологии, поведения, физиологии и пр. в линии «к человеку». Мы покажем, что это неверно: концепт «самодоместикации» объясняет лишь часть изменений3, но не движущих факторов (сходство с урбанизацией «диких» видов здесь куда эвристичней). Больше того, объяснения «самодоместикацией» ставят телегу впереди лошади, ибо объясняемые так изменения в антропогенезе ведомые, а не ведущие; последние, наоборот, изоморфны наблюдаемым в урбанизированных популяциях «диких» видов позвоночных.
У «городских» птиц и млекопитающих, также как в антропогенезе (и у собак, которым люди ставят задачу) равным образом отбираются варианты DRD4 и SERT, увеличивающие позитивное отношение к новизне, готовность устойчиво действовать в условиях новизны, вопреки сопротивлению среды, склонность к бродяжничеству, т.е. к инвазии в новую, необычную среду обитания вместо отступления от неё в спокойные и привычные условия. DRD4 кодирует дофаминовые рецепторы D2, а эта система, с одной стороны, мотивирует (система вознаграждения), с другой, поддерживает моделирование движений по образцу, увеличивает их точность и успех деятельности (запуск именно и только в момент, инициированный внешней ситуацией, чтобы успеть ею воспользоваться или уйти от её негативных последствий).
Дофамин выступает здесь как нейромодулятор, «приспосабливающий» общую схему движений к конкретной ситуации, а не только как нейромедиатор. Как показал ещё Лоренц, лучший способ повысить упорядоченность в себе и в среде вокруг — начать действовать самому, даже если дёргаться случайным образом (т.н. ауторитмия), а уж если это направленное действие на основе определённой гипотезы — тем более. Отбор вариантов DRD4, мотивирующих к подобному, способствует прогрессу познания мира, поскольку успех/неуспех действия в условиях неопределённости/новизны эту «гипотезу» поддержит или опровергнет, заставив придерживаться её альтернативы. То и другое увеличивает знание действующего субъекта о мире, толкает к ещё большему продвижению в «следующую» неопределённость и новизну, по мере того как «прежняя» сокращается в результате прошлых действия. И т.д., и т.п., почему человеческое познание мотивируется его действующим телом, у животных — наоборот, деятельность толкает их «вернуться» к отреагированию инстинктов и стереотипов, вместо продолжения и расширения познания.
Так человек может «обойти» плату за интеллектуальную деятельность, или даже превратить её в выигрыш, животные, даже антропоиды — нет.
Анализ генов, оказывающихся «под отбором» при урбанизации, с одной стороны, и при доместикации, с другой, показывает: 1) это разные гены; 2) их генные варианты, «поддерживаемые» селективным давлением в том или другом случае, связаны с разрешением существенно разных проблем приспособления. Следовательно, оба процесса «запускаются» разными стрессорами и «на выходе» создают разные адаптации. Какие именно? мы анализируем далее4.
Урбанизация меняет индивидуальность: роль DRD4 и SERT
Два кандидатных гена урбанизации (DRD4, SERT) участвуют в оптимизации выбора поведения в проблемной ситуации: задача, критически важная для «городских» птиц, «лесным» достаточно следовать видовым стереотипам. DRD4 и SERT задействуются в её двух аспектах, дополняющих друг друга: отношении к новизне, воспринимаемой как потенциальная опасность (включая способность устойчиво действовать в тих условиях), и дифференциация реальной опасности от «просто беспокойства», с ростом резистентности к последнему. «Городские» особи постоянно оказываются в обоих, а отреагирование ситуаций, разрешающее проблему или использующее её на благо животного, происходит при аномально высокой нестабильности, изменчивости и гетерогенности среды. В этом суть средового стресса в урболандшафтах, их коренное отличие от других «рукотворных», а тем более от природных [Фридман и др., 2016, рис.5].
В урбопопуляциях разных видов показан отбор вариантов DRD4 и SERT, облегчающих оба аспекта оптимизации выбора [Müller et al., 2013ab, 2014, 2017; van Dongen et al., 2015; Riyahi et al., 2015; Holtmann et al., 2016]. Данные изменения независимы в разных «городских» популяциях, образуя гомологические ряды как между ними, так и между разными видами. Направленное накопление «городских» вариантов SERT и DRD4 фиксируется не везде: в части изученных урбопопуляций оно запаздывает, подтверждая вывод о том, что при урбанизации генетические изменения сильно отстают от морфологических, а тем более эколого-поведенческих, поскольку все три связаны с адаптацией общего значения, к урбосреде как таковой, и только попутно — к отдельным существенным воздействиям в ней.
Ген SERT связывают c подавлением стресса, обычно возникающего при многозадачности, необходимости переключения между действиями, при столкновении с новым и необычным. SERT — ген серотониновой стресс-лимитации, он облегчает перенесение всякого стресса (так называемая серотониновая эйфория), в том числе при исследовании нового и необычного «вопреки» усилению разнотипного беспокойства и «возмущений» среды. Чем облегчается существование в ней на каждом этапе её освоения «дикими» видами: начальных, когда это мощный стрессор, сдерживающий рост будущей «городской» популяции везде, кроме небольшого спектра «стартовых» биотопов, и конечных, когда этот рост активно идёт, а птицы не только успешно решают проблемы «городской жизни», но и всё больше обращают её обстоятельства себе на пользу [Riyahi et al., 2015, 2017]. Это верно и для «городских» изменений гена DRD4, почему оба они суть адаптации общего значения.
На селекцию урбанизацией SERT отвечает быстрее всего. Дивергенция «городских» популяций чёрного дрозда Turdus merula от «лесных» затрагивает экзонный микросателлит, показывающий признаки положительного отбора в 10 популяциях из 12 изучавшихся (рис.2). В городских популяциях T.merula сравнительно с реликтовой частота основного аллеля снижена, а гетерозигот повышена: специфического «аллеля региона» не выявлено даже среди редких, т. е. это параллельные изменения под действием отбора [Müller et al., 2013a; Garroway, Sheldon, 2014].
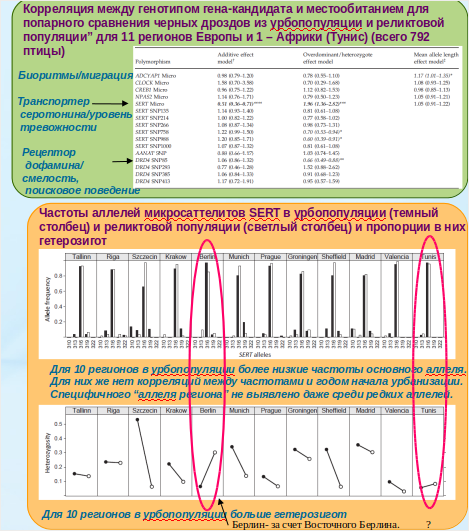
Рисунок 2. Однотипные изменения «генов урбанизации», DRD4 и SERT, в 11 городских популяциях чёрного дрозда сравнительно с «лесными» Источник. «Гены урбанизации…», op.cit.
Видимо, их «городские» аллели обеспечивают большую резистентность к тем ситуациям и объектам, на которые не надо реагировать (преобладающим в потоке событий вокруг), чем позволяют избегать ущерба и устойчиво существовать «внутри» давки и суматохи городской жизни. Благодаря им «городские» птицы «равнодушней» к последней, «сельские» при тех же воздействиях чаще всего отлетают прочь и сильно стрессируются, если остались на месте. Значительно меньше они задействованы в ситуациях, действительно связанных с «обращением Савла в Павла» — доброжелательностью к объектам, ситуациям и воздействиям, раньше вызывавшим лишь тревожно-оборонительную реакцию.
Поэтому SERT (вместе с DRD4) вносит вклад в большую смелость как у «городских» популяций сравнительно с «сельскими» и «лесными» (данные по чёрному дрозду [Müller et al., 2019], большой синице [Timm et al., 2019]), так и у популяций, освоивших новые территории за пределами основного ареала. У лесных завирушек в Новой Зеландии избирательно накапливаются варианты SERT, связанные с большей смелостью, сравнительно с британскими птицами [Holtmann et al., 2016]. У людей ген обуславливает тревожность и связанные с ней формы поведения, причем для различных аспектов тревожности влияния разных аллелей на фенотип могут быть как независимыми, так и зависимыми от локальной обстановки. На её фоне, в свою очередь, могут появляться или исчезать связи аллелей с признаками организма, определяемыми другими генами. Сама же тревожность — прямое следствие нестабильности среды [Cавостьянов и др., 2014].
Неслучайно DRD4 и SERT считают «генами индивидуальности», «биологической» подосновой выделения в популяции альтернативных стратегий поведения. Их разделение, в свою очередь, «оформляет» индивидуальность птиц, специализирующихся как «носители» этих альтернатив. Именно в городских популяциях их разделение проявляется ярче всего, как и дихотомия между «носителями»: «быстрыми» vs «медленными исследователями» у синиц, «черночеканными» сизарями vs «диким типом» и пр. [Fidler et al., 2007; Фридман, Ерёмкин, 2009].
Так, в «городских» популяциях больших синиц и черноголовых гаичек значимо больше «быстрых исследователей» («fast» explorers). Будучи помещёнными в новое пространство, они осматривают его интенсивней (быстрей движутся, чаще перемещаются), но «поверхностней» (чаще возвращаются к уже осмотренному). Им противопоставляются птицы, обследующие медленней, но много тщательней («slow» explorers). «Медленные исследователи» преобладают в исходных популяциях: «быстрые» смелей, агрессивней, но и стереотипней, склонны к копированию кормовых методов других птиц, «медленные» осторожны, робки, но инновативны, и кормятся самостоятельно. Проблемные ситуации первые разрешают активным действием, вторые — пассивным переносом (проактивная vs реактивная тактики) [Sih et al., 2004; Thompson et al., 2018].
Большая интенсивность перемещений (в том числе энергозатратных, вроде полётов и перепархиваний), увеличение порывистости каждого из них и зигзагообразности общей траектории прямо связаны с лучшей ресурсообеспеченностью в городах5, резко расширившей возможность «активного бездельничанья». Эти черты индивидуальности наследственны, как и другие анализируемые здесь: отбор по ним в 4-х поколениях больших синиц был успешен и усилил их взаимную альтернативность [Fidler et al., 2007; Riyahi et al., 2015, 2017; Baugh et al., 2017].
Скорее всего, в урбосреде важны оба варианта, но более выраженные, чем в исходных биотопах: отсюда отбор, производимый урбанизацией. У больших синиц и черноголовых гаичек постепенно вырисовывается связь альтернатив «быстрых» vs «медленных» исследователей с трейд-оффом между скоростью и точностью решения проблемных задач и задач на социальное обучение [Lefebre, Aplin, 2017]. У первых преобладает скорость, у вторых точность, почему внутри города они «сортируются» между разными элементами урболандшафта [Sprau, Dingemanse et al., 2017].
Наследуемость обоих вариантов индивидуальности также возрастает при урбанизации, в том числе потому что среди влияющих на неё генов важней всего именно SERT и DRD4. Последнее подтвердилось экспериментально. Кратковременное воздействие антагонистов рецепторов дофамина для D1 и D2 метаболических путей (последний включает рецептор D4) немедленно изменяло индивидуальность астрильдов Estrilda astrild. Блокирование D2 снизило социабельность (время, проводимое у зеркала в соответствующем тесте, и двигательную активность перед ним). Экспериментальное увеличение активности D2-пути действовало противоположно: повышало подвижность в социальном контексте (у зеркала), но снижало в других; D1 — увеличивало время, проводимое у кормушки, и интенсивность питания. Эти манипуляции также разрушили исходную корреляцию (более подвижные астрильды менее социабельны) [Silva et al., 2020]. Именно урбанизация разрушает поведенческие синдромы, присущие «сельским» особям: исчезновение связей между составляющими их качествами позволяет каждому из них меняться независимо от всех прочих при освоении данным видом данного города. Накопление «городских» вариантов обоих генов, видимо, функция времени от начала урбанизации вида и успешности её в данном городе.
Третий ген, ADCYAP1, связан с биоритмами и изучен намного меньше. Он, скорей всего, вовлечён в снижение перелётности птиц «городских» популяций, особенно урбанизирующихся «через зимовку», и/или в «раздвигание» временных границ сезона размножения с освоением урбосреды. Возможно, он не включается в урбанизацию оседлых видов и/или видов птиц вне умеренного пояса.
Влияние DRD4 и SERT на поведение неплохо изучено (в основном, правда, на людях врачами), позволяя строить предположения о функции вариантов, накапливаемых в «городских» популяциях отбором. DRD4 задействован в лучшей дифференциации ситуаций и оптимальном распределении времени между конкурирующими задачами. Он также связан с приоретизацией внимания, категоризацией происходящего (собственно лишь на людях можно количественно изучать эти характеристики), объёмом оперативной памяти, что (среди прочего) определяет участие его «городских» вариантов в усилении переключаемости и многозадачности у птиц (как и «длинных» аллелей гена сравнительно с «короткими» у человека6). Это же верно для млекопитающих. Так, «городские» койоты Canis latrans в Чикаго смелее и агрессивней, чем «сельские» (в том числе к человеку), при снижении индивидуальной повторяемости и росте контекстуальности обоих характеристик, особенно второй. В отличие от «сельских» зверей, изменения смелости и агрессии не коррелируют между собой у «городских» койотов [Wurth, 2018]. Однонуклеотидные полиморфизмы (SNP) в третьем экзоне гена DRD4 у людей и других млекопитающих связаны с межиндивидуальной изменчивостью по уровню неофилии.
DRD4 жёстче связан с чертами индивидуальности у воробьиных Passeriformes, нежели у прочих птиц (однако у ряда видов такой связи не найдено, либо, как у большой синицы, найдена лишь в ряде популяций7). В этом качестве он влияет на страх и его противоположность — стремление к новизне и исследованию. У воробьиных он и эволюционирует быстрее всего. Исследованиями индивидуально меченых особей мухоловки-белошейки Ficedula albicollis, чёрных лебедей, лесной завирушки показана связь вариантов DRD4 с межиндивидуальной вариабельностью дистанций взлёта [Garamszegi et al. 2014; van Dongen et al. 2015; Holtmann et al. 2016, обзор см. Blumstein, 2019]. Сокращение дистанций отражает резистентность к общему беспокойству в урбосреде, способность выделить в потоке событий главное и/или требующее немедленного ответа, и сразу отреагировать на него, переключив поведение без «инерции», обычной для «лесных» птиц.
Отсюда естественна связь DRD4 с урбанизацией вида. Чёрные лебеди вокруг Мельбурна значительно осторожней, чем в городе, их реакция страха и бегства гораздо инерционней, они отлетают дальше и пр.; «городские» особи у большинства видов птиц отличаются именно этим [Miranda, 2017; Фридман, 2018; Liker, 2020]. В городе они подпускали ближе и были устойчивей к беспокойству: разница связана с межпопуляционным различием генотипов DRD4 [van Dongen et al., 2015]. Связь определённых вариантов этого гена с урбанизацией уже установлена у шести видов птиц (у наполеонова бархатного ткача Euplectes afer — с созданием популяций на новых территориях птицами, улетевшими из неволи [Müller et al., 2014, 2017]). Так, в 4-х городских популяциях большой синицы (595 птиц) из Дании в отсеквенированном гене DRD4 найдено 98 однонуклеотидных замен (SNP), ассоциируемых с изменением поискового поведения. Они связаны как с кодирующими, так и с некодирующими участками, первые несут следы позитивного отбора. Наибольшая ассоциация с изменением поисковой активности — с SNP экзона 3 (особенно в 830й позиции), поменьше — с SNP экзонов 2 и 4 (рис.3). Изменения шли параллельно во всех урбопопуляциях, максимум ассоциаций отмечен в городах Вестерхейде и Ловерсмер. Там же, но не в «лесных» поселениях (Boshoek и лес Витам, в котором исследовали «сортировку» особей вдоль средовых градиентов) они связаны и с исследовательским поведением [Müller et al., 2013b].
![Рисунок 3. Изменения гена DRD4 в связи с урбанизацией больших синиц однотипны происходящим при отборе на «быстрое» vs «медленное» исследовательское поведение. В первом случае птицы взяты из Германии, во втором из Дании. Источник. Презентация «Гены урбанизации и доместикация» A.O. Брагина, И.В. Чадаева, В.В. Суслова, Ю.Л. Орлова к докладу на VII съезде Вавиловского общества генетиков и селекционеров, посвященном 100-летию кафедры генетики СпбГУ [Брагин и др., 2019].](http://www.socialcompas.com/wp-content/uploads/2022/07/Snimok-ekrana-ot-2022-07-08-08-36-50.png)
Рисунок 3. Изменения гена DRD4 в связи с урбанизацией больших синиц однотипны происходящим при отборе на «быстрое» vs «медленное» исследовательское поведение. В первом случае птицы взяты из Германии, во втором из Дании.
Источник. Презентация «Гены урбанизации и доместикация» A.O. Брагина, И.В. Чадаева, В.В. Суслова, Ю.Л. Орлова к докладу на VII съезде Вавиловского общества генетиков и селекционеров, посвященном 100-летию кафедры генетики СпбГУ [Брагин и др., 2019].
Лабораторная селекция в пользу стратегии «быстрого» или «медленного исследования» у больших синиц всего за 4 поколения меняла частоту вариантов SNP830T, в гене DRD4, но не инделя 15, взятого для контроля. То есть, разные аллели SNP830 способствуют проявлению разных типов стратегии и/или индивидуальности: С/С — второй, С/Т и Т/Т — первой. Видно доминирование «быстрого исследования» над «медленным». Сходные изменения частот данных вариантов отмечены при селекции за и против «быстрого исследования» относительно неселектированной популяции [Fidler et al., 2007]. Современный город гетерогенен настолько, что обе стратегии поведения (или же типы индивидуальности) больших синиц нашли там себе место, «сортируясь» между участками урболандшафта с разным характером беспокойства. Одновременно при урбанизации вида носители разных стратегий отбираются на большую выраженность каждой из них, вызванную в том числе отмеченным изменением гена DRD4 (о чём говорит и вышеуказанный общий рост частот полиморфизмов в третьем экзоне, и в позиции SNP830 в урбопопуляциях). Об этом говорит и отбор в лаборатории, и выявленное там полное доминирование: при неполном эта выраженность была бы ниже.
Отбор «городских» вариантов DRD4 больших синиц сопровождается и эпигенетическими изменениями. В урбопопуляциях уровень метилирования CPG-островов в его промоторе и экзоне 3 (содержит два CPG-острова, один из которых заходит в интрон 3) был значимо, хотя и не сильно, выше в образцах ДНК как из мозга, так и из крови (на ≈1%). В городских и в исходных популяциях CPG-остров в промоторе одинаково слабо метилирован (8%) относительно экзона 3 DRD4 (93%) [Riyahi et al., 2015].
Вообще, метилирование ДНК а) различается между городскими и исходными популяциями, включая локусы DRD4 и SERT; б) как минимум у больших синиц, земляных вьюрков G.fortis, G.fuliginosa и кроличьих сычей связано с поведенческими различиями между ними; в) анализ их транскриптомов показывает устойчивые различия в профилях экспрессии множества разных генов между «городом» и исходными биотопами [Riyahi et al. 2015; Verhulst et al. 2016; Watson et al., 2017; McNew et al., 2017]. Что говорит о значимости эпигенетических различий для канализации изменений, связанных с их освоением. Последнее согласуется с важностью стресса, особенно в раннем онтогенезе, для программирования изменений, повышающих устойчивость существования в будущей среде обитания [Suslov, 2013; Суслов, 2014; Роговин, 2019].
Наполеонов бархатный ткач Euplectes afer разводился в клетках. Птицы, бежавшие из неволи, с 1984 г. создали «городские» популяции в Испании, с 1992 г. в Португалии. В обоих отмечен отбор по гену DRD4 в 16 разных SNP и связь отбираемых SNP с решением проблемных задач в условиях новизны. Отловленных птиц выдерживали в клетках, после чего предъявляли два незнакомых объекта, съедобный и нет. В ответ большинство из них просто усиливало ненаправленную активность, хотя не отступало или показывало страх, как «сельские» особи. Меньшинство, напротив, снижало активность и скупо, но целенаправленно исследовало предметы визуально, либо манипулировало с ними (рис.4). Наибольшая связь с этой более сложной реакцией выявлена в Португалии у SNP449 и SNP698, в Испании — у SNP449 [Müller et al., 2014].

Рисунок 4. Урбофилы, урбофобы и “одичанты”. Источник. «Гены урбанизации…», op.cit.
Полиморфизм DRD4 влияет на восприятие новизны у птиц и млекопитающих
В случае SNP830T→С у синиц не возникло ни новой стратегии, ни комбинаторики старых. Что бывает, когда это имеет место, показывает исследование связи аллелей DRD4 («длинные» — 4 повтора vs «короткие» — 2 повтора) c избранными стратегиями карточной игры и готовностью селиться в зоне очевидного риска, близ вулкана Мерапи в Индонезии (хоть опасно, но земля плодороднее). Стратегия ва-банк (рисковый игрок: большие ставки, готов всё поставить на карту, чтобы либо выиграть, либо потерять это) коррелирует с гомозиготой по «длинным» аллелям DRD4, нерисковая стратегия (малые ставки: выигрыш невелик, но и проигрыш тоже) – с гетерозиготой. Гомозиготы по «коротким» аллелям ассоциированы с третьей стратегией игры. Игроки делают и большие, и малые ставки, размер ставки в выборке игроков варьирует8.
Вокруг вулкана Мерапи соотношение стратегий игры и генотипов выглядит как 50% нерискующих игроков (4/2 повтора), ≈23% из выборки с варьирующими ставками (2/2 повтора) и ≈27% ва-банк (4/4 повтора). Вдали от него ва-банк и нерискующих ≈по 42%, остальное – игроки с варьирующими ставками. Аллель с четырьмя повторами реже встречался в зоне риска (48%), чем вдали от вулкана (59%), с двумя наоборот (48% и 36%), остаток – минорные аллели с другим числом повторов [Faurie et al., 2016]. Видна невыгодность варьирующей ставки в Индонезии, хотя невыгодность можно снизить, если жить у вулкана. Среди поселившихся там будут и больше готовые рисковать, переехав туда (коль не родились у Мерапи), и более осторожные, готовые не задумываясь бросить всё при первых признаках извержения. Более выгодна стратегия ва-банк, хотя наиболее выгодна всё же вдали от вулкана: её приверженцы скорее поселятся там, скорее и фатально недооценят риск извержения. Резюмируя, выгоднее всего рисковать минимально, что выражено сильнее всего опять-таки у вулкана, в зоне наибольшего риска.
В отличие от синиц здесь гетерозиготы не просто демонстрируют одну из поведенческих альтернатив (как бы суммируясь фенотипически с одной из гомозигот), но связаны с новой комбинаторной стратегией9. В конкретной обстановке исследования все три стратегии «выстроились» по градиенту риска, чем «подчеркнули» неповторимость каждой из них. Эта трихотомия несводима к тривиальному дизруптивному отбору или гетерозису10. Ещё интересней была бы ситуация множественного аллелизма и/или эпистаза, особенно на фоне более разнообразного ландшафта риска, чем близ вулкана, где можно лишь уйти или остаться. Здесь каждый из >2 типов совладающего поведения может быть адаптивно ценен и сам по себе, и как ещё один фактор разнообразия такого ландшафта, меняющий ценность других типов совладевания (как при «сортировке» особей с разными поведенческими потенциями вдоль средовых градиентов).
Другие исследования млекопитающих (прежде всего верветок, собак, наконец, людей – существуют дизайны опыта для взрослых, детей, бессловесных младенцев, и этно- и геногеография) также говорят о
1) связях разнообразия аллелей DRD4 (и/или генов, работающих с DRD4) с готовностью к риску и неофилией, выбором стратегий совладевания, с успехом в комбинативных играх (а у людей и в выявлении ключевых слов), с тревожностью от встречи с новым объектом и тревожностью от повторной встречи, наконец, с поведением при решении задач: “быстрый” vs “медленный” поиск, импульсивность (помехи отвлекают от задачи) vs выдержка (решая задачу на помехи могут взять поправку), поиск удовольствия, когда и неудача, и успех ингибирует деятельность vs. погоня за удовольствием, когда и то, и другое стимулирует и т.д.; 2) локально-обстановочной селективной ценности вышеуказанного разнообразия и о сортировке и (со временем) отборе генотипов по уровням разнообразия и/или непредсказуемости среды; 3) множественном аллелизме и/или эпистазе указанного разнообразия — общий или близкий фенотип развивается параллельно: как на идентичных, так и разных генотипах; 4) указанное разнообразие никак не влияет на базовый интеллект, врождённый или выученный, в последнем случае облегчая лишь ситуативную комбинаторику и/или приоритезацию воспоминаний при поправке на обстановку, но не влияя на способность как ранее запоминать, так и сейчас вспоминать.
Антропогенез сходен с урбанизацией «диких» видов тем, что создан направленной инвазией особей во всё более экстремальную среду — сперва в саванну из лесов по градиенту их фрагментации, затем по всем средовым градиентам саванн с освоением спектра биотопов от полупустынь до побережий рек и морей. Результат — освоение всей Африки, выход из неё и поход разными путями на самую линию Уоллеса (эректусы) и за неё (сапиенсы). В обоих инвазиях психика индивидов приспосабливается к нестабильности, гетерогенности и изменчивости среды на уровне, не то, что в исходных, но порой и в недавно освоенных местообитаниях им незнакомом, то есть эволюция идёт вместе с экспансией (см.ниже). «Сортировка» и отбор среди аллелей DRD4 и комбинаторных эпистатических комплексов генов, работающих с DRD4 — логичный путь генетической адаптации при инвазиях «вверх по градиенту» экстремальности среди. Домашние животные, наоборот, эволюционируют по «изолиниям» этого градиента (охотничьи, служебные, пастушьи собаки — у какой группы пород служба экстремальнее?) или даже по нему вниз (овцы, морские свинки и т.д.). «Вверх» по градиенту домашних животных «ведёт» человек, о чём говорит эволюция DRD4 у собак [Niimi et al., 2001; Nara et al., 2005; Hejjas et al., 2007; Wan et al., 2013].
Так, у немецких овчарок в Венгрии налицо эпистаз: одни и те же аллели DRD4 дают разный фенотип на разном генетическом фоне. У псов с домашним типом поведения отсутствовала связь между активностью или импульсивностью деятельности и аллелизмом по повторам в экзоне 3 DRD4. Данную связь демонстрировали лишь псы с полицейским типом поведения из полицейского питомника и сходные с ними овчарки из Будапештского отдела этологии. Гомозиготы по «коротким» повторам (2R) показывают значимо меньшие активность и импульсивность поведения сравнительно с гетерозиготами и гомозиготами по «длинным» повторам: здесь это два варианта 3R [Hejjas et al., 2007].
Это интересно сравнить с данными по выборке финнов, отслеженных с раннего детства до 14 лет. Разница по реакции на новизну между носителями коротко- и длинноповторных аллелей DRD4 здесь фиксируется лишь в неблагоприятной среде: воспитании в строгой дисциплине и эмоциональной черствости к поведению ребёнка [Keltikangas-Järvinen et al., 2004]. У служебных собак формирование полицейского типа поведения, и вообще любого желательного, идёт как дрессурой (≈воспитанию у людей), так и прямым отбором. В пределе собака «выдаёт» породное поведение без дрессуры, как у промысловых, оленегонных и ездовых лаек, где отбор по экстерьеру до недавнего времени не интерферировал с отбором по рабочим качествам пород (кстати, и при доместикации лис).
С XIX века для многих пород рабочие качества и вообще поведении оценивались лишь при наличии определённого экстерьера, с отбраковкой отклонений от «типа». Европейский расизм, связывающий человеческие качества с особенностями морфологии, распространился и в эту сферу: считалось, что «должный» экстерьер надёжней всего обеспечивает «должное» поведение в «должных» обстоятельствах [Гийо, 2019]. Недавний анализ показывает, что собаки разных пород в целом сходны по поведению, приписываемому им людям — хозяевами и заводчиками, наследуемость межпородных различий невелика (в среднем 9%, максимум у готовности слушаться команд хозяина и общительности), большая часть изменчивости поведения создана воспитанием [Morrill et al., 2022]. У человека естественный отбор непрямой, телесность индивидов, их физиология, морфология и генетика «подгоняются» под меняющуюся социальную и когнитивную нишу, посредством общинных, национальных или государственных традиций воспитания подрастающего поколения. Сравнительно с человеком собаки и тем более большие синицы короткоживущи, поэтому у них нет свойственных нам изменений отношения к новизне (и её связи с вариантами DRD4) с возрастом или социальным статусом [Gelernter et al., 1997]. Поэтому данная связь в долговременном Балтиморском эксперименте не была установлена [Vandenbergh et al., 1997].
Множественный аллелизм гена DRD4 исследован у собак и людей. Аллелизм по повторам в экзоне 3 у сибирских хаски богаче, чем у 23 прочих пород [Niimi et al., 2001; Nara et al., 2004; Hejjas et al., 2007], что прямо связано с использованием их как ездовых. Служебных или пастушьих собак в основном отбирали на действия небольшими группами конспецификов11 или в паре с человеком, ездовые же действуют (часто и содержатся) в коллективе, отбираемом на выносливость и невозмутимость в работе. Упряжка несмотря ни на что должна продолжать бег, благоприятствует тому обстановка (что надо использовать), мешает или появилась возможность сменить задачу. Частота генотипов 2R у хаски низка, «длинных» повторов (5, 8 и гетерозиготы по ним) высока: здесь они сходны с волками, подтверждая генеалогию породы [Wan et al., 2013]. Последние не обнаружены у венгерской популяции немецких овчарок [Hejjas et al., 2007], но зато есть в японской [Ito et al., 2004].
Хотя бы один «короткий» аллель в генотипе хаски повышал оценку активности-импульсивности поведения, делал его «более домашним» в «батарее» поведенческих тестов. Вместо того, чтобы, заметив незнакомого человека, продолжать прежние действия в его присутствии, эти хаски искали контакта с ним самостоятельно и/или ориентируясь на дружелюбие хозяина. Из всей «батареи» для проблематики книги важней всего тест приветствия (англ. Greeting Test): к собаке на длинном поводке, в присутствии хозяина подходит незнакомец, дружелюбно заговаривает с ним и с собакой, гладит её. Если реакция пса дружелюбна, он отходит и пытается подозвать собаку либо проводит другие манипуляции, вплоть до взятия проб слюны [Wan et al., 2013].
Сходно тестируются поведение человеческих младенцев (приближение незнакомца→улыбка и дружелюбный разговор→попытка контакта→попытка мягкого ограничения свободы) [Lakatos et al., 2003] и поведение лисиц, норок, крыс в доместикационном эксперименте [Трапезов, 2014, 2020; Дугаткин, Трут, 2019]. Однако в первом (частью и втором) случае тестируемые имеют свободу выбора: одновременно с тестом могут в знакомой обстановке продолжать/проявлять всякое поведение из большого и обычного для них репертуара, с тестом не связанного, в разной мере отслеживая ситуацию, реагируя на незнакомца и/или пользуясь «социальными подсказками» хозяина либо взрослого у младенца.
В третьем случае свободы выбора нет. Лис и крыс тестируют прямо в клетке (месте настолько же знакомом, насколько ограничивающем репертуар поведения), норок — в клетке, примыкающей к домику, где может быть сымитирован тест открытого поля (пол расчерчен на квадраты). Скрыться в домик тестируемые норки не могут (рис.5). Они, как и «беляевские» лисы или крысы, не могут во время теста проявить (продолжать) поведение, не связанное с последним. Даже отказаться от контакта с человеком можно лишь безраздельно сосредоточившись на активном или пассивном избегании (рис.1) — от поведенческих демонстраций до отступления в угол клетки или навале на её дверцу (лисы). В обстановке, сравнимой с тестом приветствия, общаются с человеком только самые высокодоместицированные лисы — в вольере вокруг Беляевского домика и в нём самом на ферме ИЦиГ СО РАН. Животных туда помещают или они там рождаются и растут (рис.1, также см. [Дугаткин, Трут, 2019]).
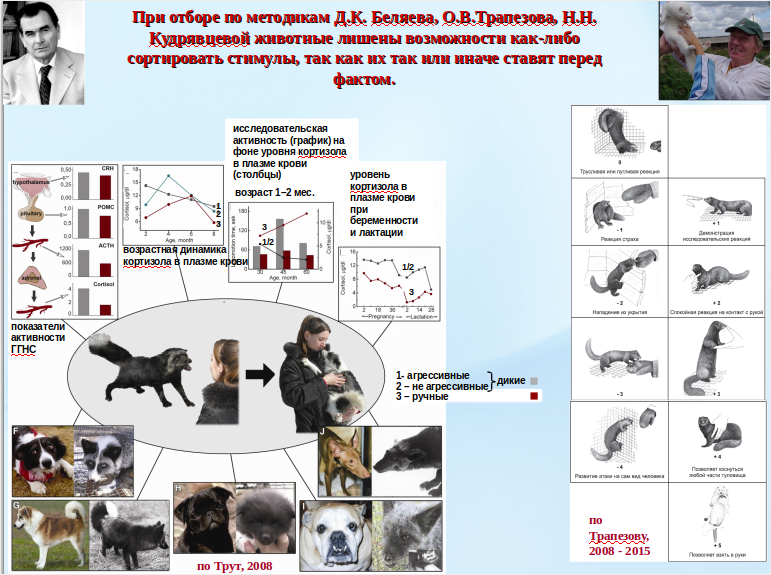
Рисунок 5. Тестирование норок на миролюбие/позитивное восприятие человека в рамках доместикационного эксперимента
Источник. «Гены урбанизации…», op.cit.
Хаски с «длинным» аллелем не игнорировали незнакомца, но и не стремились к контакту [Wan et al., 2013]. Cлужебные или ездовые собаки отбираются именно по данному поведению. Оно же предпочтительное (если не единственно возможное) для «городских» птиц, вынужденным и отслеживать новизну, и продолжать жизненно-важную деятельность на её фоне. При этом подвижность и общительность животного не страдают — ни у тестируемых хаски [Wan et al., 2013], ни у птиц. Однако хотя бы в одной (венгерской) популяции немецких овчарок отбор «сдвинул» детерминацию подобного поведения с «длинных» на более «короткие» аллели DRD4 вследствие общего обеднения аллельного репертуара экзона 3 [Hejjas et al., 2007]. Здесь налицо гомологические ряды Н.И.Вавилова между «длинными» и «короткими» аллелями DRD4 из разных популяций: внутри каждой из них те и другие «кодируют» не сходный, а фенотипически полярный признак.
Данные гомологические ряды зафиксированы и у Homo sapiens. У европейцев синдром гиперактивности-дефицита внимания (как станет понятно ниже, это компонент «быстрого» исследования, присущего также «городским» птицам) ассоциирован с нечастым, но и не редким у них 7-повторным аллелем экзона 3 (DRD4*7R), у этносов Дальнего Востока (но не его арктического сектора) — с частым там 2-повторным [Chang et al., 1996]. Среди японцев показатели поиска новизны (≈стратегии «быстрого» исследования) у гомозигот DRD4*5R значимо выше, чем у гомозигот DRD4*2R и DRD4*4R. Генотип DRD4*7R здесь не выявлен12, поэтому значимой связи не найдено при способе группировки выборки, отработанном на европейцах, где DRD4*7R нередок («короткие» аллели против «длинных»)
[Reist et al., 2007; Tsuchimine et al., 2009]. Вообще у японцев роль “европейского” DRD4*7R в поиске новизны и отношении к ней отчасти играет DRD4*5R [Тakeuchi et al., 2015], а отчасти он делит её с DRD4*2R [Chang et al., 1996]. В этносах Дальнего Востока, обладающих DRD4*7R (корейцы, филиппинцы), с ним наиболее сходен по фенотипическим проявлениям именно DRD4*2R [Reist et al., 2007].
В связи с чем показательны существенные различия аллельного репертуара у этносов Индии с их исторически высокой эндогамией и сложной историей. При общем падении частоты 7R с запада на восток, потомки традиционных индусских каст сохранили богатый репертуар аллелей (от 2R до 7R; у маратхов добавлен 9R). У мусульман же не выявлены 5R и 7R (кроме запада Индии, где частоты 7R низки), у монголоидных манипури ещё и 6R [Ghosh, Seshafri, 2005; Bhaduri et al., 2007]. У лабораторных верветок Chlorocebus aethiops с различиями в повторах DRD4 связаны разные уровни активности-импульсивности поведения и различия в отношении к новизне, как социальной (реакция на подсаженного чужака), так и новизне окружения (реакция в группе на большой незнакомый предмет). С другой стороны, есть ГР: сходное влияние на указанные характеристики поведения могут дать как типичные, так и редкие в популяции аллели DRD4 [James et al., 2007].
Cледовательно, в отношении генов DRD4, важных для устойчивого существования в экстремальной среде и/или устойчивых действий в проблемной ситуации (включая легкость переключения между разными формами активности) отбор может действовать двояко: повысить частоту аллеля, ассоциированного с адаптивным признаком, или сформировать ГР, так или иначе (изменив эпистаз, генокопировав — механизм реализации селективных давлений здесь неважен13) адаптивно “сдвинув” фенотипический признак на аллель, наиболее частый в данной популяции. Эти сдвиги возможны лишь для комплексных признаков, где субпризнаки сохраняют возможность относительно самостоятельной работы (не экспрессии!), и поэтому могут быть перекомбинированы14. Так отбор не может работать ни с химической реакцией (хотя именно положение “один ген — один фермент” удачно «легло» в СТЭ [Левонтин, 1978]), ни с эмерджентными признаками [Herron, Freeman, 2014], каждый из которых незаменим в комплексе15, ни с признаками, тесно увязанными в какой-либо синдром, баланс, адаптивный компромисс или иное ограниченное пространство возможностей16. Хотя Вавилов неоднократно подчёркивал связь своего Закона гомологических рядов (ЗГР) со свободой для комбинаторики и отбора, именно это положение на 100 лет выпало из теории гомологического сходства. Там, напротив, довлело ограничение на комбинаторику и отбор17.
Отсюда понятно, почему межэтнические сравнения не всегда однозначны, а в литературе DRD4 то связывают с поиском новизны / её исследованием / реакцией на неё, то отказывают ему в этой связи. Так, генотип DRD4*7R — предиктор высокого риска отчисления и плохой успеваемости студентов славянского происхождения Сибирского государственного медицинского университета (СибГМУ) и Томского госуниверситета. На первом курсе из них с таким генотипом было 19,4%, на пятом-шестом — лишь 3,5%, из-за отчисления. Они отличались и склонностью к аддикциям — выпивке, наркомании токсикомании. Однако у студентов финно-угорского и монголоидного происхождения (с риском быстрого развития алкоголизма) данный генотип не имеет подобных эффектов [Ильинских и др., 2015]. С другой стороны, даже в рамках одного этноса, компактно живущего, мономорфного антропологически и генетически, фиксируются устойчивые различия частот аллелей DRD4*7R в зависимости от образа жизни (при богатом репертуаре аллелей в его генофонде).
Например, они выявлены у детей ненцев, чьи родители живут в городах Ямало-Ненецкого автономного округа vs заняты кочевым оленеводством (исследование в интернатах [Ильинских, Ильинских, 2016]). Данные о сдвиге частот аллелей DRD4 в ответ на изменение хозяйственного уклада (земледелие vs традиционная охота+собирательство) у индейцев кайнганг, гуарани-ньяндева, и гуарани-кайова показывают, что такая дифференциация внутри этноса требует считанных поколений18, почему достигается скорей «сортировкой» или брачной ассортативностью, нежели отбором с его элиминацией. Птицы оказываются в «более выгодном» положении — однотипная связь DRD4 с урбанизацией выявлена у разных видов и в территориально удалённых популяциях одного вида, хотя региональные различия в ассоциированности с урбанизацией SNP экзона 3 также присутствуют.
Присущая человеку ассоциация полиморфизма экзона 3 DRD4 с реакцией на новизну свойственна также прочим приматам. Помимо собак, она зафиксирована у верветок [Bailey et al., 2007; James et al., 2007]; также гомологизирован полиморфизм DRD4 между человеком и макаком-резусом Macaca mulatta [Trefilov et al., 1999], на двух данных видах изучена связь полиморфизма DRD4 с тревожностью и импульсивностью поведения детёнышей в популяциях [Fairbanks et al., 2012; Coyne et al., 2015]. У резусов выявлена и присущая людям связь полиморфизма DRD4 с агрессивностью [Barr, Driscoll, 2014]. Сам вариабельный район в экзоне 3 распространён у разных видов обезьян и возник ещё у общего предка приматов Старого и Нового Света [Matsumoto et al., 1995; Inoue-Murayama et al., 2000].
Однако выделение каждого из изученных филумов сопряжено с утратой предковой изменчивости в этом районе и её последующим восстановлением, уже видоспецифическом, сообразно «своей» среде обитания и проблемам приспособления к ней [Livak et al., 1995]. Следовательно, сам вариабельный район в экзоне 3 и гомологичен у разных видов, однако изменчивость в нём у каждого вида своя, развившаяся параллельно: тот самый случай, когда гомологию и аналогию нельзя чётко противопоставить друг другу [Вавилов, 1935а]. С другой стороны, у крыс, как и у птиц, района с повторами нет [Gazi et al., 2000], хотя фенотипические проявления DRD4 сравнимы с таковыми у человека [Zhang et al., 2004; Li et al., 2018].
Интересней всего, что DRD4, столь сильно ассоцированный с важными человеческими качествами, от восприятия новизны и готовности рисковать до агрессивности/импульсивности поведения, не удаётся связать с модной сегодня теорией “самодоместикации человека”. Он не отмечен среди “генов доместикации”: ни по данным дифференциальной экспрессии у домашних животных, либо у животных и человека [Theofanopoulou еt al., 2017], ни по данным подобных сравнений человека с обыкновенным шимпанзе или бонобо: внимание здесь уделяется другим дофаминовым рецепторам [Babbitt et al., 2010; Marchetto et al., 2019; Staes et al., 2021]. Это значит, что либо ищут “под фонарём, а не там, где теряли”, либо фенотипические влияния DRD4 сильней обусловлены эпистазом, чем дифференциальной экспрессией. В пользу второго предположения о говорит связь с этим геном субпризнаков поиска и/или исследования новизны, но ограниченных полом. Так, когнитивная эмпатия у европеоидных женщин выше у носителей “длинных» аллелей DRD4, у мужчин — “коротких» [Uzefovsky et al., 2014].
Также ограничена полом ассоциация экзона 3 DRD4 с решением задач справедливого делёжа в игре «Ультиматум» у китайцев [Zhong et al., 2010]. Справедливость — тип совладающего поведения в проблемной ситуации, когда один игрок делит сумму денег, а второй решает, приемлемо ли деление — превысило оно некий справедливый минимум или нет19? Если нет, денег не получат оба. У рождённых зимой (с октября по март) мужчин здесь связи с полиморфизмом экзона 3 нет. У “зимних” женщин-гомозигот DRD4*4R «порог справедливости» разделения выше, чем у гомозигот 2R и гетерозигот 2R/4R, также как у “незимних” мужчин. Напротив, у “незимних” женщин-гомозигот 4R данный порог ниже гомозигот 2R и гетерозигот 2R/4R (различия на уровне тенденции).
Интересно, что ограниченные полом субпризнаки часто связаны с межиндивидуальным исследованием новизны, в субпризнаках, строго ориентированных на индивида, а не группу, связи с полом не выявлено [Görlick et al., 2015]. Видимо, у людей, в отличие от синиц, гендерное разделение и/или неравенство задаёт разные роли полов в поиске/исследовании новизны. В субпризнаках, ориентированных на группу (птенцов во время выкармливания) зависимая от пола связь разных аллелей DRD4 (знаменитый SNP830) с исследованием новизны фиксируется и у синиц. На пике выкармливания разным парам синиц цепляли к дуплянке яркий незнакомый предмет (меняя цвета, чтобы устранить привыкание), фиксируя время задержки, после которого родители решатся возобновить принос пищи птенцам. Дальше исследовали зависимость данного параметра от генных вариантов DRD4 синиц и прочих, прежде всего величины выводка и дня эксперимента. Значимая связь выявилась лишь у самцов: носители аллелей С/С делают значимо медленнее всех прочих, С/Т — быстрее, Т/Т — ещё быстрее, но между двумя последними различия не значимы (Т доминирует). Прочие факторы: порядок кормления, день опыта, размер кладки на время задержки значимо не влияют, только аллель. У самок разница между аллелями незначима, а зависимость противоположна (на уровне тенденции наблюдали падение времени задержки в ряду С/С, С/Т, Т/Т. А вот связь времени задержки с размером выводка у самок сильна, значима и отрицательна [Timm et al., 2015].
Другое возможное объяснение тех же данных — проблемы с самой дифференциальной экспрессией DRD4: не для всех эволюционно-значимых признаков она описываема в в простых усредненных рамках типа “выше-ниже”. Дифференциальность экспрессии может быть намного комбинаторно сложнее. Так, ген АSIP, задающий окрас шерсти собак, кроме аллелизма экспрессии «выше-ниже» из-за комбинаторики «мощностей» двух промоторов, также дифференциально экспрессируется на спине (циклическая экспрессия) и брюхе (сплошная) одной и той же особи, что видно по зонарности окраса волоса. К этому может добавиться перецветание окраса из-за возраста и даже погоды в данном сезоне (генетика неясна, возможна связь с регуляцией роста самого волоса) и доминантно-чёрный окрас из-за нефункциональности самого белка, см. лекцию Степана Белякина «Ветеринарная геномика».
Возможности подобной комбинаторики тем выше, чем длиннее путь от гена к признаку [Вавилов, 1935а]. И наоборот: чем сильнее этот путь связан с олигомеризацией по В.А.Догелю, или с иными ограничениями, например, физико-химического характера (актуально для животных и/ или признаков микромасштаба), тем возможностей меньше. Странно, но именно второй вариант, впервые отмеченный в Законе корреляций классиком антиэволюционизма и одновременно сравнительной анатомии Ж.Кювье [Cuvier, 1805] стал магистральным путём развития эволюционных теорий (!) ХХ, ныне и ХХI века. Закону корреляций Кювье равно соответствуют основные концепты всех них: гомеостат Шмальгаузена, ключевое изменение Г.Ф.Осборна, адаптивный компромис А.П.Расницына, или случайная эволюция по Е.В.Кунину20. В такой эпистеме эволюция возможна только кювьерианским манером — от одной катастрофы (разрушения корреляций) до другой.
Однако сравнив человека с другими бипедальными формами, видим, что никакой катастрофы в антропогенезе не происходило. У птиц, динозавров, приматов (сифака, гиббоны, шимпанзе), грызунов (тушканчики), сумчатых (кенгуру) передние конечности исключались из ходьбы/бега так, что всё равно давали с ногами кювьерианские корреляции — через осанку или другие органы (например, питания — у карнозавров, муравьедов, наземных ленивцев, возможно, копытных-халикотериев: кстати, налицо гомологический ряд Вавилова, если за признаки считать сами корреляции). В линии развития от австралопитеков к Homo sapiens нет ничего подобного21: вместо этого человеческая рука развила возможность функциональной комбинаторики здесь и сейчас с практически любым внешним органом, в том числе ногами, где руки могут помогать равновесию, осанке и походке (скажем, у оррорина Orrorin tugenensis), хотя могут этого и не делать. Прибегнув к математической аналогии, можно сказать, что в антропогенезе эволюция шла на рост функционально-комбинаторной мощности множества, включающего руки и внешние органы индивида — вместо установления скоррелированности их «работы» (тогда мощность в лучшем случае не растёт).
Сразу отметим: в литературе нами не найдено прямых ассоциаций полиморфизма DRD4 с таким важным для доместикации и ключевым для антропогенеза субпризнаком поиска/исследования новизны, как совместное внимание (joint attention), поиск “социальных подсказок” и совместное действие на их основе [своего рода “кооперативность мышления”, Hare et al., 2012; Хейр, Вудс, 2014]. Есть указания лишь на непрямые ассоциации [Gangi et al., 2016], и не только с рецепторами дофамина [Stavropoulos, Carver, 2013]. При множестве работ по ассоциации DRD4 с реакцией младенца на незнакомца практически нет экспериментов сходного дизайна с влиянием на упреждающую улыбку младенца — часть совместного внимания. Другие же составляющие последнего, вроде взаимоотношений мать-детёныш [Coyne et al., 2015, Fairbanks et al., 2012]) ассоциированы не только с DRD4, но и с фоновым обучением — у беспокойных матерей дети беспокойнее. Показана связь данного признака с другими обстоятельствами, «рождающими» модификации: сезоном рождения, метилированием, дистрессом матери во время беременности [Zohsel et al., 2014]. Всё это создаёт возможность генокопирования, т. е. увязывания субпризнаков в жесткий, всё более и более постоянно действующий, автономный от прочих признаков синдром, а не комбинаторное расширение общей генетической основы признаков при локализации экспрессии каждого.
Поиск и исследование новизны строятся совершенно иначе. Здесь успех сам по себе «ликвидирует»: отрицательная обратная связь если и есть, то по исчерпанию продукта и/или времени, тем более просто отсутствие информации — тоже новизна, но в форме неизвестности. Отсюда, исследование новизны не может идти постоянно и автономно: это локально-обстановочная функция организма, не его гомеостаза или онтогенеза, изученных Г.Турессоном и И.И.Шмальгаузеном в контексте генокопирования. Тогда поиск новизны совместным вниманием, и поиск новизны животным-доместикантом равно неидентичны DRD4-асоциированому поиску новизны: cобаке и беляевской лисице должно быть скучно не с хозяином, а без него, этого самого ненового человека! Идентичны соображения для совместного внимания. Всё это важно для сравнения (и противопоставления друг другу) когнитивных «достижений», связанных с урбанизацией или доместикацией.
Мы видим, что сама по себе биохимическая реакция рецептора DRD4 с G-белком, за что отвечает экзон 3, как и всякая параллельная ей биохимия [Hodgson et al., 2019], лишь вносит вклад в поведенческие признаки, связанные с поиском новизны, её исследованием и совладанием22, однако ещё не определяет их! Прибегнув к математической аналогии, можно сказать, что биохимический вклад не может быть элиминирован по модулю, см. данные по нокаутам, однако по вектору изменяем отбором практически до противоположного23: от “сдвига” фенотипа на чуть менее длинные повторы, выявленного у собаки и человека (с запада на восток в Старом Свете24), до обнуления и инверсии фенотипического эффекта DRD4*7R — снижение интереса к новизне вместо его роста, как у американских негров с аддикцией или «белых» американок [Gelernter et al., 1997].
Такой сдвиг возможен, постольку
а) под отбор попадает весь комбинаторный комплекс признаков, связанных с динамикой дофамина в синапсе при передаче сигнала — независимо от того, что их кодирует25,
б) постольку, поскольку такой отбор вообще идёт, то есть для популяций актуален как выбор организмом окружающей его здесь и сейчас обстановки, так и выбор стратегий совладевания. Скажем, невозмутимость = помехоустойчивости поведения, реализуемого особью равно несмотря на его собственный выбор обстановки или его отсутствие (она оказалась там, куда её поместил человек или непреодолимые силы стихии).
Актуальность обоих выборов или лишь одного (обстановки или совладания) может быть ограничена попаданием популяции в однообразную среду (выбора нет) или отбором в пользу оптимизации выбора в среде разнообразной26.
Кроме того, всякая автономизация от среды ставит оба выбора в отношение взаимного исключения друг друга, тогда как «городским» птицам и млекопитающим важна комбинаторика выборов, их амальгама. При доместикации, наоборот, нет выбора обcтановки и крайне ограничен выбор совладающего поведения: по факту оба выбора предоставляет Беляевский домик (рис.1), однако попасть туда пока могут только уже высоко задоместицированные лисицы или их потомство. Отбор на оптимизацию (увы, один из ведущих трендов в современной теоретической биологии) этих выборов в любой их комбинации равносилен ловушке эндемизма. Он держит популяцию в исторически типичной среде, где только и возможно поведение “дикого типа”, то есть блокирует урбанизацию.
Нокаут же DRD4, например, у мышей, ведёт только к комплексному снижению неофилии и росту безусловного страха, не влияя на общую активность [Keck et al., 2013] и условнорефлекторный страх [Falzone et al., 2002]. Выполнение сложных двигательных задач здесь даже улучшилось [Rubinstein et al., 1997], если мышь вообще решается действовать — она больше сидит в укрытии [Dulawa et al., 1999]. Двигательная активность растёт, но лишь потому, что подобная мышь стремится сбежать в укрытие [Dulawa et al., 1999] или хотя бы избегать центра в тесте открытого поля [Keck et al., 2013]: базовый интеллект не нарушен, но проявить его можно только там, где оба выбора организму неактуальны27. Также у человека базовый интеллект не нарушен независимо от того, какие у него аллели DRD4 [Nöthen et al., 1994; Roussos et al., 2009]. В крайнем случае (ежедневные сеансы длительного иммобилизационного стресса) исследовательская активность мышей-нокаутов даже росла, но нарушалась её когнитивная обработка, что напоминало шизофрению [Tan et al., 2019]. Поэтому de facto мощность данной активности не росла.
Иными словами, DRD4 не связан непосредственно с интеллектом — способностью особи к решению задач «умственным» или двигательным усилием. За что же он тогда отвечает? Как сплавить в амальгаму неофилию и невозмутимость, рисковость и осторожность, если абстрагироваться от интеллекта? И как соединить с ними ассоциацию у человека полиморфизма DRD4 с болезнью Альцгеймера и шизофренией (расстройство памяти и внимания); сексуальными, ментальными и посттравматическими расстройствами, рисками аддикций (избирательная память на фоне расстройства функции подкрепления поведения) и агрессивностью, тревожностью, синдромом гиперактивности-дефицита внимания (расстройство и того и другого); способностью к обучению, освоению языков, а у птиц к распознаванию песен (взаимоотношения избирательной и общей памяти) и прочему, описанному в литературе?
Логичный вывод из такого набора взаимоисключающих признаков — DRD4 отвечает за приоритезацию внимания [Görlick et al., 2015]. При посредстве DRD4 все рецептируемые организмом здесь и сейчас объекты подразделяются на
I) Важные здесь и сейчас. Внимание к ним повышено, их исследование, или иное эмоционально окрашенное отношение к ним (например, избегание по прошлому жизненному опыту) в мозгу особи подкрепляется в первую очередь,
II) Неважные здесь и сейчас. Внимание к ним ослаблено, поведение относительно них слабо подкреплено: они не игнорируются, но нет и стремления к контакту или иной связи с ними.
III) фоновые.
О фоне организм если и помнит, то лишь чтобы на нём отслеживать изменения в категориях I и II, и оценить нужность пересмотра всех категорий здесь и сейчас. В фон могут попасть жизненно важные объекты, но здесь и сейчас их приравнивают к самым пустячным, а в следующий момент изменение остановки переведёт их в I. Если вспомнить военно-мемуарную литературу («Живые и мёртвые» К.М.Симонова, «Огонь» А.Барбюса), то приоритезации соответствует поведение в бою [Драгомиров, 2012], особенно рутина действий бойцов на фронте. За многодневной канонадой им повседневное поведение просто нельзя отложить, в отличие от ожесточенного, но относительно короткого боя. Бывалый солдат продолжает рутинные дела, ибо отличает опасную канонаду от пальбы неопасной — по соседней позиции, с неверным прицелом и т.д. (хотя от ошибок не застрахован), а под фоновую просто спит, зато просыпается от тишины.
Полководец же профессионально обязан приоритезировать на поле боя28 [Драгомиров, 2012], меняя ради этого даже длительность боевой обстановки. Иначе его место в лучшем случае в отделе планирования штаба, см. Г.К. Жуков [2002] о маршале С.К.Тимошенко. Это не менее верно для следопыта, см. Дерсу Узала в пургу на озере Ханка [Арсеньев, 2020]. Отсюда следует разница между фоном и помехами, не всегда отслеживаемая даже в литературе по кибернетике29 [Корогодин, Корогодина, 2012]: фон принадлежит к данной обстановке, помехи к ней не принадлежат, они из другой обстановки. Фон может быть и не важен, но полезен, помехи важны лишь в отрицательном смысле — чем их больше, тем больше мешают. Возможность превращать помехи в фон резко увеличивает потенциал комбинаторики и эволюции30, тогда как ЭТЭ и номогенез в лучшем случае говорят о помехоустойчивости и обнейтраливании помех, а СТЭ просто приравнивает их к случаю.
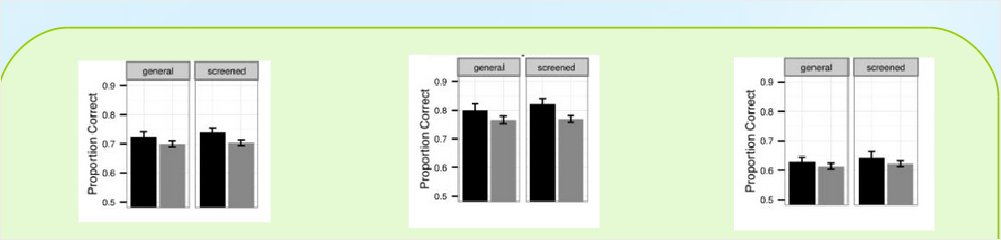
Тогда приоритезация связана с умением не столько классифицировать, сколько переклассифицировать. Дизайн опытов для проверки этих выводов отработан пока только для человека [Görlick et al., 2015]. В задаче обучения категоризации (category learning task) используют фигурки: у каждой 10 качественных признаков, у каждого признака — одно из двух альтернативных состояний (рис.6). Испытуемым из выборки, полиморфной по DRD431, предъявляют две фигурки-прототипа, состояния всех признаков которых взаимно альтернативны. Задача испытуемого — указать, к какому из прототипов будут принадлежать фигурки, которые ему ещё предъявят. Для её решения каждому испытуемому случайно указывают категорию — один признак прототипа32, по состоянию которого надо классифицировать принадлежность фигурок к прототипу (рис.6). Сперва испытуемые проходили предварительное обучение, затем решали задачу, что и оценивалось. В изученной выборке носители «длинных» аллелей DRD4 (7 и более повторов) лучше решали задачу, чем носители «коротких» (6 и менее) повторов. Различия между ними (рис.7) значимы по данным ANOVA 2 (screened, general) × 2 («длинные» DRD4, «короткие» DRD4), особенно в выборке с улучшенным предварительным обучением (Screened: её участники прошли батарею тестов на нейропсихологическое здоровье: отсутствие аддикций, синдрома дефицита внимания и гиперактивности, черепно-мозговых травм и др.) [Görlick et al., 2015].
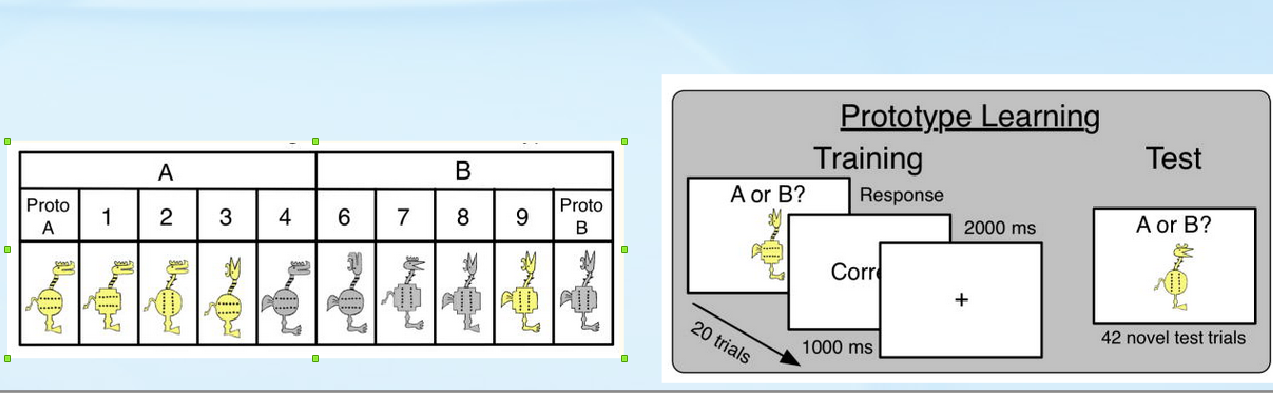
Рисунок 6. Признаки объектов, используемых в задачах на категоризацию, обучение по прототипам и тестирование в работе Görlick et al., 2015.
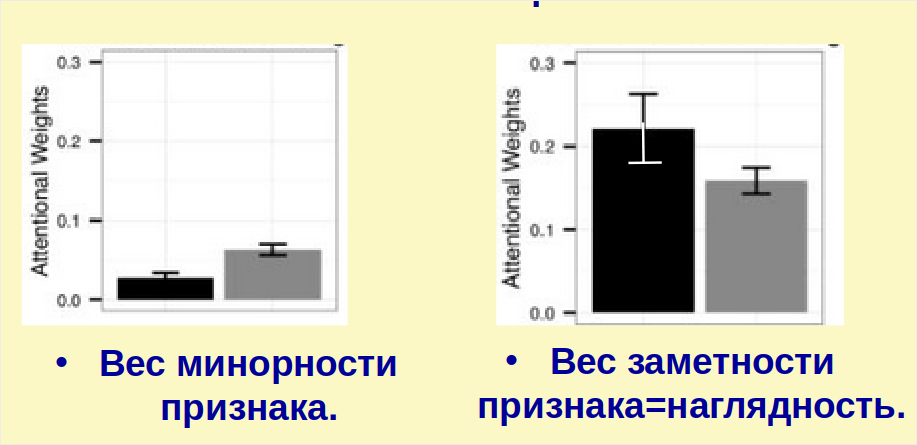
Хотя каждый из 10 признаков одинаково важен для решения задачи, некоторые были заметней для испытуемых. Как и ожидалось, на правильность категоризации влияют схожесть с прототипом, контраст и количество признаков с отличающимися состояниями. Похожие на прототипы фигурки категоризуются лучше, чем несхожие, фигурки с отличием у 1-2 признаков лучше, чем 3-433. В опыте удалось оценить вес минорности признака (отклонение опыта от компьютерной категоризации, где все признаки имеют априорно равный вес), характеризующий априорное представление испытуемых о главных (голова, туловище, цвет) и второстепенных (минорных) признаках фигурки, и заметность признака (чем больше испытуемых игнорирует этот признак, тем выше этот вес) (рис.7).
По мере решения задачи внимание испытуемых поляризуется, но лишь у носителей «длинных» аллелей DRD4. Это значимо повышает точность категоризации: минорные признаки делаются ещё минорнее, но те, что оказались заметными — ещё заметнее [Görlick et al., 2015]. Идентичные результаты получены в опытах по распознаванию речи: «длинные» аллели улучшали распознавание на фоне информационных, но не энергетических помех [Xie et al., 2015]. Информационные аудиопомехи этой работы соответствуют визуальной задаче приоритезации в: [Görlick et al., 2015], ибо толкают на неверное неправильное отнесение компонентов шума к целевому высказыванию, и наоборот, отвлекают внимания от последнего и т. п. из-за того, что несколько людей говорят одновременно о разном34.
Везде критически важен не поиск целостного образа, а всё более скорое сосредоточение внимания на небольшом числе признаков35 [Görlick et al., 2015] с возможностью их реконструкции, пусть частичной, но осмысленной (идентификация ключевых слов — не ясно, что делал данный субъект, но говорят именно о нём, неясно, кто бежал, но говорят о бегущих и т.п.) [Xie et al., 2015]. Разница между носителями «длинных» и «коротких» аллелей сохраняется также при контроле импульсивности поведения [Görlick et al., 2015]36, и в выборке Screened. В отличие от опытов с индонезийцами [Faurie et al., 2016], здесь первые доминировали над вторыми [Görlick et al., 2015; Xie et al., 2015].
I.1) а), б), в)

д) е)
I.2) a) б) в)
г) д)

II.
Рисунок 7. Статистический анализ результатов решения задач 1) на категоризацию (I. Выборка General. II. Выборка Screened), 2) на объём оперативной памяти (i — общий объём, ii — пропорция верно воспроизведённых букв).
Обозначения. Темный столбик – гомо и гетерозиготы по длинным аллелям, светлый столбик – гомозиготы по коротким аллелям. а)-д) — см.текст. Из: [Görlick et al., 2015].
Отсюда вырисовывается связь «длинных» аллелей DRD4 с синдромом гиперактивности-дефицита внимания [Görlick et al., 2015]. Его носители просто не обращают внимания на черты, которые считают скучными и/или неважными, и тем сильнее, чем эти черты константнее. Это освобождает время и энергию для тех занятий, которые им же расценены как важные или легко выполнимые здесь и сейчас. Однако, переклассификация в дизайне задачи на обучение категоризации анизотропна [Görlick et al., 2015] или усечена [Xie et al., 2015]: есть поляризация внимания, но не проверено умение переоценить признаки — изменить их важность сообразно моменту, локальной обстановке (или проверено a posteriori).
Выяснилось, что и с этой проблемой у европеоидов лучше справляются носители «длинных» аллелей. Тестирование младенцев даёт позволяет отвлечься от профессиональных навыков, интеллектуальных, личных, культурно-языковых37 и прочих пристрастий в минорности и заметности признаков. Так, взрослые белые американцы с DRD4*7R — наибольшие индивидуалисты, японцы с DRD4*2R — наибольшие коллективисты, то есть ярче носителей иных аллелей выражают особенности психологии культуры, присущие данному обществу в целом [Kitayama et al., 2014]. Младенцы с «длинными» DRD4 более активны в ситуации свободной игры, но менее гневливы при легком физическом удержании [Auerbach et al., 2001] и менее тревожны при появлении и постепенном приближении незнакомца [Lakatos et al., 2003].
Однако они дольше не отвечали на его ласковую улыбку, дольше игнорировали предложенную им игрушку — длинно-, но не короткоповторный младенец продолжал свои дела, хотя и не игнорировал незнакомца [Lakatos et al., 2003; De Luca et al., 2003]. Низкий страх незнакомца при «длинных» аллелях, высокий — «коротких» сохранялся у детей и в 6 лет [Pappa et al., 2014]. Здесь очевидны как сходство этих данных с поведением служебных собак или «городских» птиц, так и различия в интерпретации: [de Luca et al., 2003] считают младенцев с «длинным» DRD4 более робкими, с худшей исследовательской реакцией в ответ на новизну, более сдержанными в выражении в ответ на неё как счастья, так и печали либо гнева, почему сделан вывод об их худшей приспособляемости. Вывод справедлив, если рассматривать его с точки зрения исключительной важности (если не единственной возможности) для младенца именно коллективного исследования новизны: скажем, совместное внимание критично для быстрого овладения языком38.
Отсюда, даже исследование знакомых вещей, но в коллективе более привлекательно, чем незнакомых в одиночку, в пределе такое исследование идёт с матерью (или другими старшими, заботящимися о детях). То есть с самым неновым для ребёнка человеком, из-за чего, несмотря на высокую новизну (=незнакомость) обстановки мощность новизны может быть низкой. Для ребёнка незнакомец одновременно и фактор новизны, и кандидат в члены коллектива. Ситуация птиц «городских» популяций диаметрально противоположная: им важно устойчиво продолжать жизненно необходимые повседневные действия (за ребёнка эту рутину делают старшие, за домашних животных во многом — сам человек) в мощных по изменчивости обстановках, всякая облигатность в комбинаторике им гибельна39. Сравнивая дизайн опытов с людьми [Lakatos et al., 2003, De Luca et al., 2003] и животными [Coyne et al., 2015, Keck et al., 2013, Fairbanks et al., 2012, Fidler et al., 2007, Bailey et al., 2007, James et al., 2007, Dulava et al., 1999] надо обращать внимание как на разные субпризнаки в общей генетической основе признака “поиск/исследование новизны”, так и на возможности комбинаторики между ними. В пользу этого говорят связь DRD4*7R c дезорганизацией поведения детской привязанности у европейцев [Lakatos et al., 2000, 2002] и описание гомозиготного носителя нефункционального DRD4 без каких-либо психиатрических патологий [Nöthen et al., 1994].
Когнитивные и эмоциональные составляющие большей смелости в «городских» популяциях птиц сравнительно с исходными уже анализировались нами: это отнюдь не смелость как таковая (пренебрежение реальной опасностью), а её лучшая дифференциация от беспокойства, большая готовность действовать несмотря на последнее. Однако в дизайне обсуждаемых опытов [Görlick et al., 2015; Xie et al., 2015] нет и намёка на опасность. У крыс воспоминания о страхе блокируются на время активации DRD1 в префронтальной коре. Побочный результат этой настоящей («слепой») храбрости — отсутствие усиления вознаграждающих эффектов у опиоидов [Li et al., 2018]40. Напротив, активация DRD4 в той же коре контролирует приобретение и выраженность воспоминаний о страхе [Li et al., 2018]: не помехоустойчивость, а “спокойное самообладание на поле боя”, важное условие практической приоритезации [Драгомиров, 2012]. Правда, обстановочный фон здесь внутренний, а не внешний.
Зато с активацией DRD4 усиливаются и вознаграждающие эффекты опиоидов [Li et al., 2018]. Данная связь с аддикцией ярко локально-адаптивна при ограниченном доступе к веществам-аддиктам, как бывает чаще всего — вспомним фронтовые сто грамм (хотя это не опиоиды). В противоположном случае эффект дезадаптивен [Ильинских и др., 2015]: носители соответствующих аллелей DRD4 могут повернуть и успехи в приоритезации [Görlick et al., 2015], и «спокойное самообладание» [Li et al., 2018] к поиску вещества-аддикта [McGeary, 2009] задолго до появления физиологической зависимости. Например, монреальские дети 4-12 лет, обладающие 7R, даже при прочих равных едят больше, считая по калорийности, что особенно выражено, когда в зоне их пешей доступности (3 км) в ритейлах низка доля полезных продуктов. Они наиболее уязвимы к развитию ожирения, хотя связь с индексом массы тела не выявлена (Paquet et al., 2021). Тем более, что у европейцев с «длинным» аллелем DRD4 кроме роста неофилии также падают зависимости от вознаграждения и от вреда избранной стратегии действий (в эксперименте это азартная игра, долгое участие в которой с большими ставками вело к проигрышу41).
Здесь также снижаются общая тревожность, аффективная пугливость и реакция на безусловные аверсивные стимулы (хотя вижу, что страшно, но не боюсь) при сохранности обработки эмоциональных стимулов в случае быстрого затухания самой эмоциональной реакции. Больше того, носители «длинных» аллелей проявляют тревожность и испуг лишь на начальное предъявление посторонних стимулов (приятных, неприятных и нейтральных), а потом их снижают, сохраняя различие приятных и неприятных, что интерпретируется как выделение помех. В этой же ситуации европейцы с «коротким» аллелем тревожность тоже снижают, но продолжают реагировать на все три типа стимулов при меньшем затухании эмоциональных реакций. Поэтому в совокупности они будут тревожнее, пугливее, менее эмоционально выносливы42: лимит времени на любое дело у них тем напряжённей, чем изменчивее среда [Roussos et al., 2010] — как “дикие” виды птиц на первых шагах освоения урбосреды. Младенцы-гомозиготы DRD4*4R также выражают свои эмоции (счастье, гнев или печаль) куда энергичней, чем гетерозиготы DRD4*4/7R [De Luca et al., 2003]. Даже большее время, затраченное носителями «длинных» аллелей на планирование решения задачи может (хотя и не обязано) сыграть им на пользу в изменчивой среде — так больше шансов для корректировки, больше мощность комбинаторики. «Короткие» повторы DRD4 выигрывают постольку, поскольку среда стабильна. Хотя общая эффективность решения задач теми и другими сравнима, вклады планирования и воплощения у обоих различны [Roussos et al., 2010].
Снижение зависимости от вознаграждений и от вреда позволяет носителям соответствующих аллелей DRD4 чаще выбирать рискованные стратегии поведения, и их дольше держаться. Они рисковые игроки и продолжают играть, несмотря на слабый/редкий выигрыш или даже на проигрыш [Roussos et al., 2010]. Однако нечувствительность к вознаграждению — ангедоничность — может заставить и искать его чаще. Вместо поиска удовольствия, когда с достижением того прекращается и поведенческая деятельность, идёт погоня за удовольствием, когда его достижение не останавливает, а стимулирует либо чаще повторять данное поведение, либо всячески его разнообразить — ибо базовый интеллект не нарушен — комбинировать, играть во множество разных игр так, чтобы вред/проигрыш одной возмещался удовольствием другой43. Действительно, европеоиды с «коротким» DRD4 характеризуются высоким уровнем возбуждения. Отсюда, подобные люди склонны избегать дополнительной стимуляции, даже если оценивают ситуацию положительно, внешне выражая счастье. Напротив, носители «длинных» DRD4 с их низким уровнем возбуждения склонны искать стимуляцию [De Luca et al., 2003]. К этому мы вернёмся в главе 5, когда будем говорить об использовании «городскими» птицами стрессоров, признанных ими не опасностью, а лишь беспокойством.
Способность к категоризации, приоритезации и их пересмотру требует памяти. Причём это кратковременная исполнительная память, «удерживающая» именно высокоприоритетные черты несмотря на решение отвлекающей второстепенной задачи, а не кратковременная рабочая (отвечающая, например, за вспоминание событий по порядку44). Первая лучше у носителей «длинных» аллелей45, причём они доминируют46, на вторую влияние числа повторов DRD4 не выявлено [Görlick et al., 2015, Хie et al., 2015]. На избирательное запоминание контекстуально новых или необычных признаков также влияет однонуклеотидный полиморфизм -521C/T в промоторе DRD4: C-аллель улучшает память, но при повторном предъявлении тех же признаков эффект пропадает [Strange et al., 2014]. Наконец, несмотря на ангедоничность, эмоциональный фон управляет приоритезацией: когда у испытуемых с «длинными» DRD4 провоцировали грустное настроение, у них росло внимание к чертам выражения лица, связанным с грустью и/или страхом [Wells et al., 2013]. Надо ли говорить, что в экстерьере носителей разных аллелей DRD4 — людей (разных рас), собак, обезьян — всеми цитированными авторами отнюдь не отмечено синдрома дестабилизации, неизменно возникающего при одомашнивании [Трут, 2007, 2008]? (как он отсутствует и у «городских» птиц).
В приоритезации и связанных с ним субпризнаках вклад DRD4 определяется внутренней и внешней комбинаторикой. Внутренняя — комбинаторика полиморфизмов разных районов гена DRD4, см. рис.7 для синиц [Fidler et al., 2007]. У человека найдено 5 таких функционально-значимых и полиморфных районов [Paterson et al., 1999]: выше обсуждавшийся экзон 3, где полиморфизм не ограничен лишь повторами; интрон 2; экзон 1, у H.sapiens ассоциированный с бредовыми растройствами [Catalano et al., 1993], и два участка в 5’-районе47. Три из них перекрывают «подпись» приспособления к урбосреде в DRD4 P.major [Müller et al., 2013b]; перекрытия в экзоне 3 [Müller et al., 2013a, 2014] и промоторе [Riyahi et al., 2015] описаны для других урбанизированных видов. Два района функционально изучены у собак [Hejjas et al., 2007, Niimi et al., 2001, Ito et al., 2004]; четыре есть у приматов [James et al., 2007, Bailey et al., 2007, Shimada et al., 2004; Seaman et al., 2000; Inoue-Murayama et al., 2000, Livak et al., 1995]. У человека, но не антропоидов, выявлен полиморфизм по тандемной дупликации в 5’-районе на 1,2 тыс. пар нуклеотидов выше инициирующего кодона. Полиморфный район охватывает сайты транскрипционых факторов [Seaman et al., 1999].
У большинства видов пока изучен полиморфизм экзона 3. Внутренняя комбинаторика даёт большое число аллелей [Lakatos et al., 2002, Ito et al., 2004; Hejjas et al., 2009], частично функционально перекрывающихся, т.к. фенотипический эффект полиморфных районов опирается на разные биохимические модули: работу с G-белком [Ma et al., 2010], метилирование [Hodgson et al., 2019], альтернативный промотор [Hejjas et al., 2009], модуляцию мощности основного промотора [Strange et al., 2014, Seaman et al., 1999] и др. Это обеспечивает гомологические ряды Вавилова — сходные фенотипы «на выходе» при разной комбинации аллелей по одним субпризнакам и различные по другим.
Внешняя комбинаторика связывает DRD4 с генами ферментов катаболизма MAOA, ALDH3A1, COMT (возможными кандидатами в «гены урбанизации»48) и с генами транспортёров моноаминов, обеспечивающими их обратный захват (SERT) [Richards et al., 2016; Iofrida et al., 2014; Alfimova et al., 2014; Barr, Driscoll, 2014; Ma et al., 2010; Armbruster et al., 2009; Auerbach et al., 2010]. Из генов синтеза отмечена функциональная связь DRD4 с геном ТН (тирозингидроксилазы) [Wan et al., 2013]. В компаунде порождаются ГР к фенотипам внутренней комбинаторики DRD4 либо фенотипы поиска/иследования новизны, которые не даёт ни один ген в отдельности [Zenner et al., 1998; Kühn et al., 1999; Schmidt et al., 2007; Lakatos et al., 2003, Wan et al., 2013]. У генов катаболизма дофамина также повышена скорость эволюции в сравнении с генами его синтеза [Ma et al., 2010]. В свою очередь, сами гены внешней комбинаторики DRD4 имеют собственную, внутреннюю комбинаторику [Kubinyi et al., 2012; Maddox et al., 2017]. Как и в случае с DRD4, значимость последней разнообразие обстановок может нивелировать или подчёркивать [Madrid et al., 2018].
В итоге, DRD4 стал самым быстро эволюционирующим из DRD2-подобных рецепторов [Ma et al., 2010] и одним из самых — среди генов рецепторов нейротрансмиттеров у млекопитающих, как в “дальнем» филогенезе (между человеком и грызунами [Iwama, Gojobori, 2002]), так и в «ближнем», наиболее быстром филогенезе («городские» vs исходные популяции птиц). Однако DRD4 нет среди генов доместикации, которым присуща высокая дифференциальная экспрессия у «ручных» или агрессивных лисиц [Wang et al. 2018] либо крыс! (доместикационый эксперимент — другой случай «ближнего» и быстрого филогенеза; масштаб филогенетического расстояния между грызунами и хищными сравним с таковым между ними же и приматами). Среди них нет и гена SERT, чьё влияние на поведение различно при противопоставлении свободы выбора поведения, разнообразия обстановок для действия особи [Maddox et al., 2017; Madrid et al., 2018] отсутствию возможности действовать, сокращению выбора, локально-обстановочной бедности (то есть именно средовым изменениям при доместикации). Сокращение возможностей выбора и действия в отношении других особей создаётся и опытом повторяющихся побед в агрессивных конфликтах, или хронической депрессии у тех, кто их раз за разом проигрывает [Кудрявцева, 2013]. Нет DRD4 и в наборе быстро эволюционирующих генов, общем для четырёх видов домашних животных и человека (также называемых «генами доместикации» ([Theofanopoulou et al., 2017]). Зато экзон 1 DRD4, cвязанного с агрессией и импульсивностью [Barr, Driscoll, 2014; Wush, 2018], попал под отбор на агресcивность у шамо — японских бойцовых петухов, и долгокукарекающих (тоже проблема с подкреплением?) петухов наганакидори [Komiyama et al., 2014].

Бойцовые петухи шамо
Подведём итог. Высокие показатели приоритезации и связанной с ней оперативной памяти делают непредсказуемую урбосреду относительно комфортной: как минимум, носители данных качеств лучше справятся с «городскими» проблемами. И наоборот: особи с низкими избирательностью внимания/объемом оперативной памяти «расценят» урбосреду как некомфортную и/или опасную, почему покинут город либо «отступят» на крупнейшие «острова» природных территорий. А это интенсифицирует «сортировку» особей между обоими ландшафтными альтернативами, ускорит отделение «городских» популяций, их дивергенцию по экологии/поведению, с одновременным снижением груза элиминации. Так микроэволюция ускоряется, и тем больше, чем значительней альтернативность обоих ландшафтов и экстремальней для вида урбосреда [Alberti et al., 2017; Johnston, Munshi-South, 2017; Liker, 2020]. Одновременно последняя делается безопасней в плане риска вымирания новообразованной популяции.
Приоритезация и оперативная память критически важны:
I. При жизни в изменчивой среде;
II. Когда формы демонстраций, используемых в коммуникации между особями — знаки (а то и символы), а не просто стимулы, пусть даже специфические. Т. е. они распознаются по инварианту формы, характерный образ которого «кодирует» значение сигнала, почему при демонстрировании воспроизводится устойчиво и точно, несмотря на «помехи», сильно увеличивающиеся в урбосреде [Фридман, 2013]. Для птиц это значимо в силу постоянства сигнального репертуара (набора таких демонстраций, используемых в охране территории, поиске партнёра, ухаживании за ним в паре и других значимых для вида контекстах общения), для людей — при базовом владении одним и тем же языком. Это же верно для сигналов среды (включая предвестники, в том числе телодвижения особей других видов) и распознавания по ним значимых для птиц ситуаций («на кормушке», «кошка поблизости», «люди гуляют», «подростки носятся», «машины едут» и пр.), с возможным их изменением в будущем. Так, бристольские чайки трёх разных видов (озёрная, клуша L.fuscus и серебристая L.argentatus) целенаправленно собираются у школ во время перемен, когда дети перекусывают на свежем воздухе. Возле центров по сбору мусора они концентрируются в часы подвоза свежих отходов, но в городском парке, где кормятся дождевыми червями и прочими беспозвоночными, активность не зависит от доступности антропогенных кормов [Spelt et al., 2020].
Урбанизированные серебристые чайки на побережье графства Корнуолл (Великобритания) боятся прямого человеческого взгляда и значимо ближе подпускают людей, смотрящих вниз, не на них. Большинство чаек бросает стянутые у людей чипсы, если на них посмотреть. 19 из 74 чаек, однако, были достаточно смелы, чтобы подойти и схватить еду, когда люди на них смотрят, хотя это заняло в среднем больше времени [Goumas et al., 2019]. Однако это не простой страх, влекущий за собой избегание: направление взгляда и действий они отслеживают, предпочитая потроганную человеком еду. «Предпочтение потроганного» распространяется и на несъедобные объекты [Goumas et al., 2020].
III. В поиске наиболее референтного носителя всех названных сигналов при взаимодействии животного с разными типами компаньонов (особи своего или близких видов, разные категории событий вокруг, значимых и не очень), а также III.1. когда нужно освоить чужую речь (на уровне смысла, а не грамматики), выявить кто здесь главный, расширить умвельт;
IV. При оценке опасности среды.
Однако ситуации I-IV заведомо отсутствовали в отборе ручных vs агрессивных лисиц или крыс/мышей в ИЦиГ СО РАН! Поэтому там до сих пор нет моделей, позволяющих изучать ситуации I-IV — как и любые другие, предполагающие гетерогенность среды и свободу выбора особью более благоприятных для неё вариантов средовой мозаики. Отбор по DRD4 в опытах Д.К.Беляева и его последователей просто не велся: не позволял дизайн эксперимента. Это хорошо объясняет отрицательные результаты анализа дифференциальной экспрессии генов по данным чип-сека [Брагин и др., 2019]. Отбор «городских» вариантов генов урбанизации не отмечен в доместикационных изменениях, изученных в ИЦиГ, в том числе у «ручных» vs «агрессивных» лис и крыс-пасюков, а также неселектированных лабораторных мышей с индивидуальным опытом побед и поражений.
Лис, как и крыс, в течении нескольких поколений отбирали на агрессивное vs толерантное поведение по отношению к человеку. Мышам лишь создали историю повторяющихся побед и поражений в стычках при ссаживаниях. Для этого к более сильной особи подсаживали заведомо более слабую — или наоборот, давали им драться по 5 минут, после чего повторяли; и так до тех пор, пока не сформируется устойчивый, но не наследуемый комплекс поведения победителя (наглый, задиристый, легковозбудимый, поэтому охотно лезущий в драку) и проигравшего (неофобный, всего боящийся, пассивно избегающий драк)49. Лисы и крысы здесь демонстрировали наследственные изменения модели поведения в ответ на стресс, мыши модификационные, но также устойчивые. Для обоих «характеров» крыс и мышей (победители vs побеждённые) изготовляли чип-секи, выявившие дифференцированно экспрессирующиеся гены в мозгу, и среди них искали «гены урбанизации» SERT, DRD4, ADCYAP1 [Брагин и др., 2019].
Для проведения анализа WGCNA на данных RNA-seq в образцах агрессивных и ручных крыс количество ридов, выравненных на ген, было подсчитано с помощью функции featureCounts в пакете Subread. Из выборки исключены гены, на которые не выравнялись прочтения RNA-seq. В дальнейший анализ вошло 18000 генов. Анализ дифференциальной экспрессии осуществлялся с помощью пакета DESeq2 (пороговое значение 0.01). Дифференциальная экспрессия между образцами считалась для трех моделей: 1) генотип; 2) генотип+отдел мозга; 3) отдел мозга.
Результат был неожидан и прямо противоречил исходным предположениям авторов. Среди топ-10 и топ-20 списка дифференциально экспрессирующихся генов беляевских крыс «генов урбанизации» вообще не было (рис.8). В первом случае идентифицировали 37 генов, чья экспрессия выше у ручных крыс и 40 — у агрессивных; не выявили и каких-либо ассоциативных связей (Genemania, ANDVisio) DRD4 с подобными генами. Из них у мышей-победителей или «проигравших» фиксируется только SERT в топ-20, но это не показательно. Он облегчает любой стресс, почему экспрессия SERT равно повышена у обоих типов мышей сравнительно с контролем, т. е. нет дифференциальных изменений. Анализ представленности терминов Gene Ontology и путей KEGG с помощью пакета limma показал, что у ручных крыс преобладают гены, участвующие в метаболизме сахаров, у агрессивных – в метаболизме пуринов и действующие при окислительном стрессе [Брагин и др., 2019].
Рисунок 8. А. Наиболее дифференциально экспрессирующиеся гены между агрессивными и ручными крысами; Б. Значимые термины Gene Ontology, экспрессия которых значимо выше либо у агрессивных, либо у ручных крыс.
Поэтому даже удивительно, что он не попал под отбор у беляевских крыс. Эксперимент на мышах показывает очень широкую норму реакции его экспрессии, уровень продукта этого гена легко поднять и надолго оставить высоким: всё перечисленное благоприятствует генокопированию. Зато в обоих моделях вместо DRD4 очень хорошо дифференциально экспрессируется DRD2. Это контринтуитивно: у больших синиц разница спектров SNP по гену DRD4 фиксируется уже на четвёртом поколении селекции50. Она, правда, шла на поисковое поведение [Fidler et al., 2007], но агрессивные vs ручные крысы беляевской линии резко различны и по нему тоже, а селектируются намного дольше. История с SERT столь же контринтуитивна.
Дальше по данным к статье Wang et al. [2018] второй автор специально искал пересечение генов, вовлечённых в урбанизацию «диких» видов птиц, с генами, связанными с доместикацией собственно беляевских лис. И не обнаружил его! Точней, если оно и есть, то в неидентифицированных областях типа ENSCAFG00000032444, которые не с чем сравнить. Общая картина у лис, видимо, такая же, что у крыс – 146 дифференцированно экспрессирующихся генов у доместицированных и диких лис отвечают за общую активность гормональных и нейротрансмиттерных систем, за регуляцию онтогенеза, но не решение конкретных проблемных задач (в противоположность «генам индивидуальности»). Их пересечений с «доместикационными» списками дифференциально экспрессирующихся генов пока не найдено: из просмотренных 20 генов нет ни одного ни в «лисьем» списке, ни в «крысином» [Сhadaeva et al., 2018; Брагин и др., 2019]. Интересно посмотреть и гены, связанные с пением у птиц, где также задействованы рецепторы DRD (у людей связанные с распознаванием нюансировки речи).
Как все это объяснить – пока не известно. Единственно, что можно сказать точно: это ещё один факт, показывающий, что Д.К.Беляев открыл яркий, красивый, интересный, но все-таки очень специфичный отбор, связанный со строительством «второй природы» человеческой деятельностью, отчасти с самим антропогенезом, и именно потому вряд ли возможный в дикой природе [Suslov, 2018; Брагин и др., 2019]. Поэтому и не получается объяснить происходящее с «дикими» видами птиц, млекопитающих, рептилий при освоении городов и других «рукотворных ландшафтов» на основе доместикационной модели. Попытки так делать — следствие нынешней научной моды — грешат постоянной подгонкой «некрасивых фактов под красивую теорию51», и всё время используют ложные аналогии, негодные для прогноза происходящего.
Есть и более общие соображения, почему нельзя прямо экстраполировать результаты Д.К.Беляева на природную эволюцию. Доместиканты, «продукты» дестабилизирующего отбора, дают яркие гомологические ряды изменения морфологии, поведения, гормонального статуса. Однако в законе гомологических рядов Н.Н.Вавилова редкие варианты признака в центрах происхождения доминантны, в опытах же Д.К.Беляева – рецессивны или полудоминантны. Как правило, особи выживают посредством доминантных генов и вопреки скрытой изменчивости любой природы (полудоминантным и рецессивным генам). Если последняя даст полезный фенотип – отбор сделает все, чтобы она перестала быть скрытой.
Во-вторых, и это главное, гомологические ряды Вавилова равно выявляются и у доместицированных, и у диких популяций/видов/родов, гомологические ряды Беляева – только у доместикантов. Последнее не зависит от таксономического статуса: одомашненные лиса, собака, корова и лошадь показывают множественные сходства между собой (в том числе и по генам, оказывающимся под отбором), у диких же псовых нет признаков собак52 и беляевских лис. Если только не считать таким признаком банальную разницу в размерах (волк и кустарниковая собака), но тут классическое селекционистское объяснение СТЭ просто бросается в глаза [Suslov, 2018; Брагин и др., 2019].
Так как DRD4 оказался связанным с приоритезацией внимания и объемом рабочей памяти, было бы интересно посмотреть гены у людей, могущие решать сходные задачи: влияющие на объем рабочей памяти, на задачи категоризации, на задачи языковой беглости (когда нужно назвать за ограниченное время термины одной категории – животных, профессии и пр.) и на решение задач на фоне отвлекающих действий. Увы, все они недоступны для применяемых в ИЦиГ методик тестирования на агрессивность/доместикантность.
Антропогенез как всё лучшее использование новизны во всё более экстремальной среде обитания
Рассказывая, почему концепция «самодоместикации» неприменима к бонобо, мы использовали среди прочей аргументации филогенетическую. Бонобо архаичней Pan troglodytes по морфологии, особенностям онтогенеза, «конструкции» социальной системы. Первый ближе к общему предку рода Pan и «ветви», ведущей к австралопитекам с людьми, второй вид продвинутей и моложе [Трут, 2008; Diogo et al., 2018; Fruth, Hohmann, 2018]. Но во всех прочих парах в таблице доместицированный вид — потомок «дикого», не наоборот!
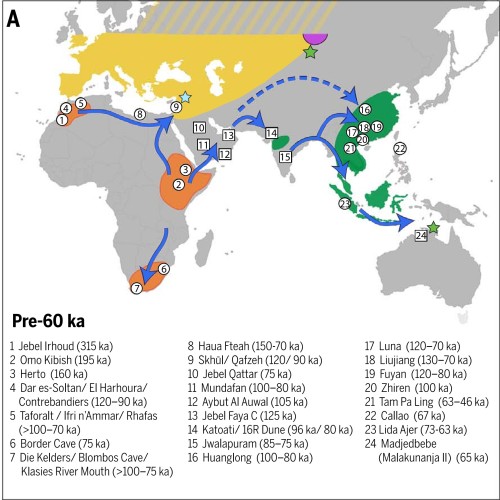
А. Карта мест находок костных останков, относимых к сапиенсам (цифры в кружочках) и археологических следов сапиенсов без костных останков (цифры в квадратиках), датировки предшествуют времени 60 тыс. лет назад. Стрелки указывают пути древних миграций (120-60 тыс. лет назад). Желтым цветом обозначен ареал неандертальцев, оранжевым – ареал ранних сапиенсов, фиолотовым – денисовцев, зеленым – ареал ранних сапиенсов в Юго-Восточной Азии. Голубая звездочка указывает место метисации сапиенсов с неандертальцами, зеленые звездочки — места метисации сапиенсов с денисовцами. Источник Генофонд.рф, op.cit.
Рассмотрим аргумент «от филогенеза» подробней. Современные и особенно палеонтологические данные говорят, что все африканские антропоиды сем. Pongidae — эндемики. Число их ископаемых находок исчезающее мало как по сравнению с гоминоидами53 (даже более примитивными, чем Homo erectus), так и с общими предками линий «к человеку» и «к современным понгидам» — дриопитеками Dryopitecus spp. [Grehan, Schwarz, 2009]. Единственный антропоид, хоть как-то сравнимый с ними по числу находок и по экологическому разнообразию их местонахождений — орангутан Pongo spp. Он же наиболее архаичный, наименее скелетно засиндромленный54 и самый эврибионтный из ныне здравствующих антропоидов.
В сравнении с ними он же самый лабильный по социальной структуре (может жить в одиночку, сохранять парные отношения и даже формировать группы), а биотопически — самый непоседливый бродяга [Delgado, an Schaik, 2010; Krützen et al., 2011]. Он же приспосабливается к антропогенным нарушениям лесного покрова вырубками и плантациями масличной пальмы (если они невелики), мирится с травянистыми пространствами [Spehar et al., 2018].
Можно сказать, что оранги настолько близки к линии австралопитеки→люди по «естественной истории» (включая ряд важных моментов генетики), насколько далеки по “родословному дереву” [Grehan, Schwartz, 2009]. Разделение двух видов шимпанзе было запущено изменением русла Конго, тогда как для дифференциации разных форм орангов55 понадобились морские проливы, созданные общей трансгрессией и катастрофами типа взрыва Кракатау.
С другой стороны, оранг ближе всего к общему предку антропоидов и гоминоидов, будучи одновременно неожиданно близок к человеку, но не к африканским антропоидам по организации проксимальных промоторов генов, экспрессируемых в мозгу (высокая пластичность регуляции, больше открытых промоторов). Как уже говорилось, DRD4 нет среди генов с высокой дифференциальной экспрессией между человеком и шимпанзе [Babbitt et al., 2010; Marchetto et al., 2019; Staes et al., 2021]. Зато DRD4 африканских антропоидов несёт общую делецию в первом (консервативном) повторе экзона 3; поэтому у человека и орангов Pongo spp. он ближе к общему предку человекообразных [Livak et al., 1995]. Широкое распространение ископаемых находок орангов на континенте говорит, что они, как минимум, пересекали открытые ландшафты между лесами. Данное поведение реконструируется и для австралопитеков, иначе не объяснить их широкое расселение по Африке.
Африканские антропоиды в лучшем случае обитатели опушки, ночующие в лесу. Реконструкция же ардипитека даёт примата менее древесного облика, чем шимпанзе, но вполне выводимого из дриопитеков, давших адаптивную радиацию в различные ландшафты. У человека надёжно установлена связь между склонностью к миграциям, в том числе хозяйственным, и полиморфизмом в экзоне 3 DRD4 [Gören, 2016]: благодаря им человек снова и снова «вводит себя» в новую и непривычную среду, часто — более или менее экстремальную, как «дикие» виды позвоночных в ходе урбанизации.
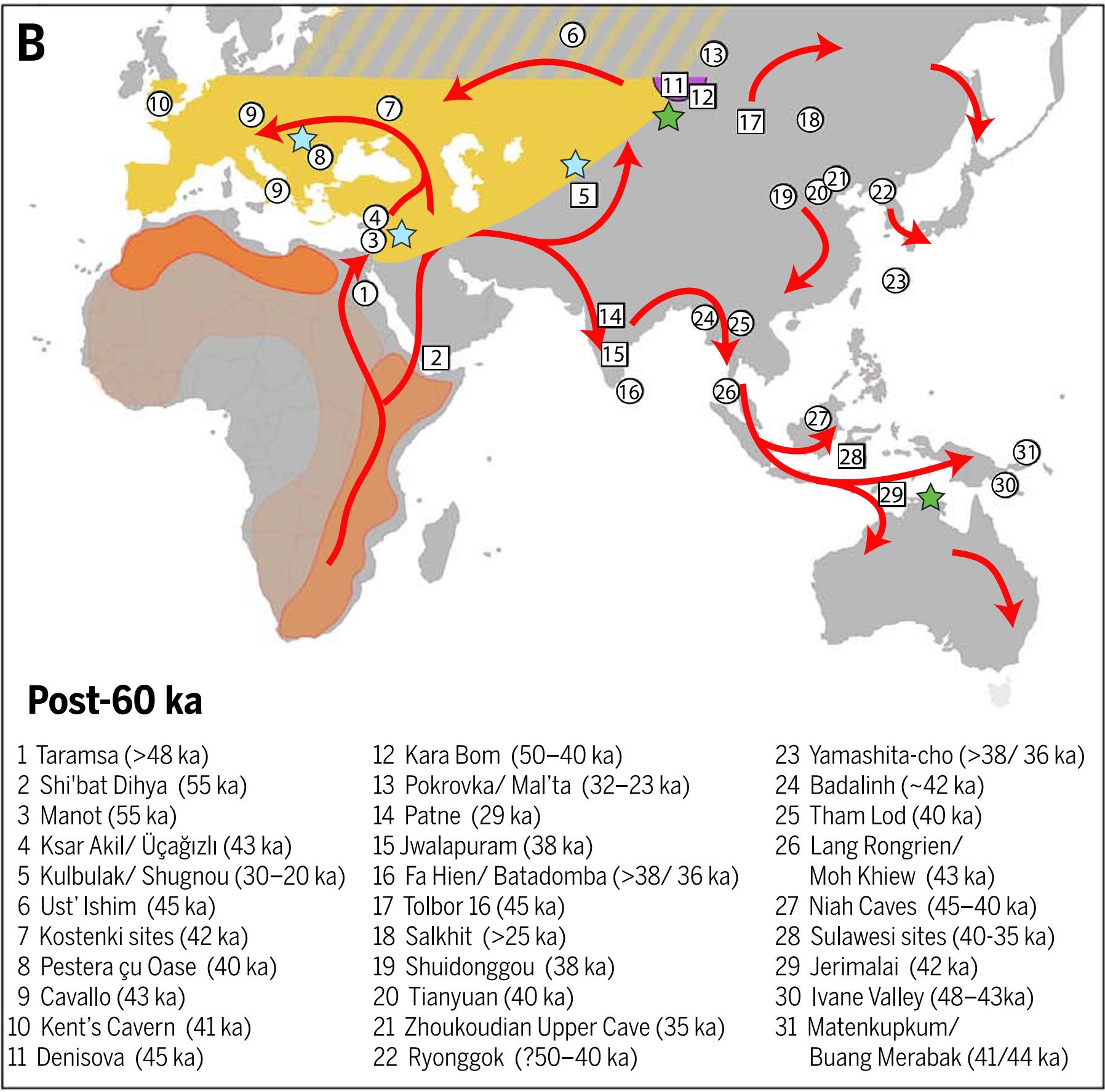
В. Карта мест находок костных останков, относимых к сапиенсам (цифры в кружочках) и археологических следов сапиенсов без костных останков (цифры в квадратиках), датировки после 60 тыс. лет назад. Стрелки указывают пути миграций (60-30 тыс. лет назад). Желтым цветом обозначен ареал неандертальцев, оранжевым – ареал ранних сапиенсов. Голубые звездочки указывают места метисации сапиенсов с неандертальцами, зеленые звездочки — места метисации сапиенсов с денисовцами. Источник Генофонд.рф, op.cit.
Реконструкция истории полиморфизмов в экзоне 3 даёт следующую картину [Wang et al., 2004]: предковый аллель 4R, 7R возникает из него в субпопуляции, чьи потомки покинут Африку, 2R — результат рекомбинации 4R/7R гетерозиготы, возможно, параллельной в разных обществах [Reist et al., 2007].
Вышеописанные затруднения легко разрешимы предположением, что общий предок человека и африканских антропоидов долгое время существовал как вавиловский вид-система из эврибионтных лесо-саванновых и стенобионтных лесных популяций, разделённых по градиентам риска и мозаичности биотопов, но время от времени скрещивающихся [Patterson et al., 2006]. Косвенный довод «за» данное предположение — сильно большие возраста остатков австралопитеков в Южной Африке, чем это полагали раньше, они одновременны кениантропу, восточноафриканским афарским и поздним анамским авторалопитекам. Это здорово увеличивает разнообразие одновременно живущих двуногих человекообезьян, живших на континенте 3-4 млн.лет назад.
Эврибионты, приспособившиеся к мозаике лесных фрагментов потом и к открытым травянистым ландшафтам с отдельными куртинами деревьев и кустарников, сохранили и предковый вариант повтора. В открытом ландшафте, как в городе, надо уметь нормально жить на фоне раздражителей, издали дифференцируя их на опасные и неопасные, пусть и беспокоящие. Последних, конечно, в саванне меньше, чем в городе, но и убежищ там меньше56. Стенобионты, всё более и более отступавшие вглубь лесов по мере их фрагментации из-за иссушения климата [Bonnefille, 2010], получили делецию, затем восстановили в собственной популяции длинные и короткие аллели DRD4, затем отделили новый вид — гориллу, а при окончательной дивергенции с лесо-саванновыми популяциями дали шимпанзе57.
Дальнейшая дивергенция афроантропоидов, равно как и дивергенции в линии австралопитеки→люди связана с морфо- и цитологической дифференциацией: ни DRD4, ни SERT не имеют на них выхода, в чем может быть ещё одна причина, почему эволюция по этим генам не выходит за рамки микроэволюции. Другой важный момент антропогенеза: в своей эволюции человек испытал значительное увеличение мозга, отличающегося от такового антропоидов не столько самими генами, сколько их регуляцией. Её пластичность в линии «к человеку» растёт, «к шимпанзе» — уменьшается: функционирование мозга человека меньше зависит от генов и сильнее меняется под влиянием среды, в том числе дольше сохраняет «детский» период высокой пластичности [Gunbin et al., 2018].
Всё вместе это реконструирует следующую картину: сперва существует вавиловский центр разнообразия дриопитеков вокруг Средиземного моря (если частично не на его дне), порождающий как резидентные, так и мигрирующие формы с ГР между ними. Дальше мигранты оттуда (в том числе в Азию и вглубь Африки) формируют в последней вторичный центр, где окончательно дивергируют гоминоиды и африканские антропоиды. Вторые дали резидентные формы, первые развили общепредковый биотопически непоседливый образ жизни, заселяя обстановки, где лесные стации чередовались с саванновыми, т. е. самые фрагментированные и нестабильные участки исходных ландшафтов.
Позже, когда климат стал засушливым, африканские антропоиды окончательно отступили в тропические леса (отсюда редкость находок). Гоминоиды же расселились «вверх» по градиенту смены леса саванной, породив множество параллельных ветвей, «обеспечивших» много находок при широкой географии. Выживали они за счет комбинаторики поведенческого разнообразия сперва между ветвями, а потом и внутри отдельных, самых эврибионтных ветвей. Поэтому анатомически гоминоиды ближе всего к шимпанзе, но экологически сперва дают параллель другой миграционно активной и одновременно самой архаичной понгидной ветви, а затем многократно превосходят её в адаптациях к дальним миграциям (доминирование прямохождения) и разнообразию поведенческих стратегий.
Наконец, размер мозга при одомашнивании уменьшается: миниатюризация возможна благодаря «кооперативности познания» и роли «социальных подсказок» в решении проблемных задач58 [Hare et al., 2012, tabl.1; Дугаткин, Трут, 2019]. Отсюда сомнения в применимости концепции «самоодомашнивания» к антропогенезу: в филогенетической линии, ведущей к Homo sapiens «напрямую», через гейдельбергского человека Homo heidelbergensis (без учёта «боковой ветви» к неандертальцу H.neanderthalensis c денисовцем H.denisovan) размер мозга, средний и относительный, возрастает, и лишь с раннего верхнего палеолита (27-25 тыс.лет назад) уменьшается59. Сомнительно её применение и к бонобо: его мозг относительно шимпанзе не уменьшен.
Однако существенный довод «за»: сравнение геномов человека разумного, неандертальца и денисовца показывает, что только у первого шёл интенсивный отбор по генам, связанным с миграцией клеток нервного гребня — стержневого процесса в доместикации, и сильно влияющего на формирование лица [Theofanopoulou et al., 2017]. Поэтому если самодоместикация и шла, то не на начальном этапе антропогенеза (дивергенция афроантропоидной и гоминоидной линий), а на самом финальном (отделение Homo sapiens от сестринских видов). В таком случае отдельные элементы самодоместикации бонобо — параллелизм эндемичной линии, диахронный к человеку разумному. В обоих случаях признаки «одомашненности» так и не «сложились» в синдром: с точки зрения «самодоместикации» его отсутствие необъяснимо.
Синдром проявлений доместикации образуется быстро и «бьёт» по размножению, как и облигатное прямохождение (однако от него не отказывается ни одна линия гоминоидов, хотя все они и без того виды малоплодные). Прямохождение ставит наш вид перед выбором «синдром доместикации или хозяйственно важный признак», постоянном сопровождающим одомашнивание животных. И тогда люди раз за разом выбирали второе, заменяя доместикацию60 отловом и приручением, полувольным содержанием, а то и просто периодическим отловом без приручения (альпак и лошадей стригли и отпускали, как делают и сейчас, разводя оленей на панты, а страусов на перья). Однако как совместить подобные практики (синдрома так и не уничтожившие, «звёздочка» на лбу обычна у лошадей, с вислоухостью собак борются жесткой выбраковкой и/или купируя уши) с самодоместикацией далеко не многочисленного, но зато широко расселявшегося человечества хотя бы на заре становления H.sapiens?
Заключение
Отсюда возможен следующий ответ на вопрос «что общего между урбанизацией и доместикацией»? В обоих случаях изменения связаны с одной и той же проблемой приспособления — к экстремальной среде, в силу разных причин «наступающей» на исходные местообитания вида, дробящей их, трансформирующей с краёв и изменяющей изнутри, в режиме медленной катастрофы, и с одним и тем же — контринтуитивным ответом на происходящее: инвазией в экстремальную среду обитания, у освоения которой есть свои (и значительные) плюсы, и мобилизация всех сил и возможностей организма для сохранения устойчивой деятельности в экстремальной среде, т. е. в условиях новизны, при необходимости лучшей классификации новых, потенциально опасных ситуаций, лучшего отделения реальной опасности от просто беспокойства, лучшего использования новых возможностей, которые даёт среда, а для этого их надо отделять от всей прочей новизны и пр.
Это ведёт к неожиданному изоморфизму изменений поведения и когнитивных способностей и психики при урбанизации «диких» видов и в антропогенезе, в подоснове которого — отбор одинаковых вариантов DRD4, лучше поддерживающих когнитивные усилия и продолжения прежней деятельности в условиях новизны и потенциальных опасностей — но и возможностей — которыми изобилует осваиваемая среда обитания (экстремальная в первую очередь в силу исторической нетипичности).
Литература
Арсеньев В.К. Встречи в тайге. М.: АСТ, 2022. 132 с.
Арсеньев В.К. По Уссурийскому краю. М.: изд-во РуДа, 2020. 432 с.
Брагин А.О., Чадаева И.В., Суслов В.В. и др. Урбанизация и доместикация// VII Съезд ВОГиС, посвящ. 100-летию каф. генетики СПбГУ, и ассоц. симп. СПб, 2019. С.695-695.
Гегель Г.Ф.В.Г. Эстетика. В 4-х тт. М.: изд-во «Искусство», 1968-1973. 330, 326, 623, 667 с.
Гийо Д. Люди и собаки. М.: НЛО, 2019. 328 с.
Вавилов Н.И. Иммунитет растений к инфекционным заболеваниям. М.: Тип. Рябушинских, 1919. 240 с.
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. Саратов, Губполиграфотдел, 1920. 16 с. Цит. по: Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. Л.: Наука, 1987. С. 10-22.
Вавилов Н.И. Географическая локализация генов пшениц на земном шаре. Л.: Издательство Академии Наук СССР, 1929a. 6 с.
Вавилов Н.И. Проблема происхождения культурных растений в современном понимании // Достижения и перспективы в области прикладной ботаники, генетики и селекции. Л.: Изд-во ВИПБиНК и ГИОА, 1929б. С. 11—22.
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. М.-Л.: Сельхозгиз, 1935а. 56 с. Цит. по: Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. Л.: Наука, 1987. С. 56-100.
Вавилов Н.И. Учение об иммунитете растений к инфекционным заболеваниям (Применительно к запросам селекции) // Теоретические основы селекции растений. Т.1. М.-Л., 1935б. С.893-990.
Вавилов Н.И. Пути советской растениеводческой пауки. (Ответ критикам)// Соц. реконстр. с.-х. 1936. №12. С.33—46.
Вёльфлин Г. Классическое искусство. Введение в изучение итальянского Возрождения. Спб.: Алетейя, 1999. 328 с.
Драгомиров М.И. Разбор романа «Война и Мир» с военной точки зрения. М. Либроком. 2012. 144 с.
Дугаткин Л., Трут Л.Н. Как приручить лису (и превратить её в собаку)? Сибирский эволюционный эксперимент. М.: Альпина Нон-фикшн, 2019. 290 с.
Жуков Г.К. Воспоминания и размышления. Т. 1. М.: ОЛМА-ПРЕСС, 2002. 415 с.
Ильинских Н.Н. Полиморфизм гена DRD4 у детей кочевых и урбанизированных коренных жителей севера Западной Сибири / Межд. научно-исслед. ж. 2017. № 12 (54). Ч.1. С.25-27.
Ильинских Н.Н., Ильинских Е.Н., Ильинских И.Н. и др. Роль полиморфизма гена DRD4 дофаминэргической системы в успеваемости и поведенческих стереотипах этнически различных когорт студентов// J. Sib. Med. Sci. 2015. №6. С.64-73.
Койре А. Очерки истории философской мысли: о влиянии философских концепций на развитие научных теорий. М.: Прогресс, 1985. 286 с.
Корбетт Дж. Кумаонские людоеды. М.: Армада-пресс. Зелёная серия. 208 c.
Корогодин В.И., Корогодина В.Л. Информация как феномен жизни // Корогодин В.И. Феномен жизни. Т.2. М., 2012. С.173-338.
Креславский А.Г. Новый взгляд на адаптивную природу полиморфизма. Концепция псевдонейтральных мутаций// Журн. общ. биол. 1993. Т.54. №6. С. 645-658.
Кудрявцева Н.Н. Нейробиология агрессии. Мыши и люди. Новосибирск. Наука-Центр. 2013. 272 с.
Кунин Е. Логика случая. О природе и происхождении биологической эволюции. М.: Центрполиграф, 2017. 582 с.
Левонтин Р. Генетические основы эволюции. М.: Мир, 1978. 351 с.
Махров А.А., Болотов И.Н. Экологические причины высокой морфологической пластичности представителей таксона, обитающих в центре его происхождения (на примере благородных лососей Salmo) // Изв. РАН. Сер. Биол. 2019. №1. С.43–51.
Противовоздушная оборона Берлина в период Второй мировой войны. М.: Воениздат, 1947. 317 с.
Раутиан А.С. Палеонтология как источник сведений о закономерностях и факторах эволюции. // Современная палеонтология. Ред. Меннер В.В., Макридин В.П. М.: Недра, 1988. Т.2. С. 76–118.
Раутиан Г.С. Гомологические ряды в окраске птиц // Природа. 1987. № 10. С.66– 74.
Роговин К.А. Стресс в раннем онтогенезе как адаптивное явление// Журн.общ.биол. 2019. Т.80. №2. С.95-123.
Савостьянов А.Н., Науменко В.С., Синякова Н.А. и др. Взаимосвязь уровня тревожности с полиморфными вариантами гена серотонинового транспортера у русских и тувинцев// Вавил. журн. генет. и селекц. 2014. Т.18. №4/3. С.1268-1280.
Северцов А.С., 2008. Эволюционный стазис и микроэволюция. М.: Т-во научн.изд. КМК. 176 с.
Синская Е.Н. Динамика вида. Л.: Сельхозгиз, 1948. 266 с.
Суслов В.В. Основное противоречие адаптации и пути его разрешения// Развитие жизни в процессе абиотических изменений на Земле. Иркутск: Байкальский музей Иркутского НЦ СО РАН (Листвянка), 2014. №3. С.102-103.
Суслов В.В., Пономаренко М.П., Рассказов Д.А. Закон или правила гомологических рядов? I. Теории эволюции и закон Bавилова. II. Ряды Копа и Вавилова // Экология и эволюция: новые горизонты. Мат-лы межд.симп. к 100летию акад.С.С.Шварца. Екатеринбург, Гуманитарн.ун-т, 2019. С.344-350.
Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. Краткий очерк теории эволюции. М.: Наука, 1977. 297 с.
Трапезов О.В. Генетические основы доместикации// Молекулярная и прикладная генетика. 2014. Т.18. С.118-132.
Трапезов О.В. Д.К. Беляев о генетико-эволюционных аспектах стресса и стрессируемости //Кролиководство и звероводство. 2020. Т.2. №. 5. С.19-22.
Трут Л.Н. Доместикация животных в историческом процессе и в эксперименте// Инф.вестн.ВОГИС. 2007. Т.11. №2. С.273-289.
Трут Л.Н. Эволюционные идеи Д.К. Беляева как концептуальный мост между биологией, социологией и медициной// Вавил. журн. генет. и селекц. 2008. Т.12. №1-2. С.7-18.
Филипченко Ю.А. Изменчивость и методы ее изучения. М.-Л.: Госиздат, 1929. 275 с.
Фридман В.С. От стимула к символу. Сигналы в коммуникации позвоночных. В 2-х ч. М.: URSS, 2013. 560 c., 424 c.
Фридман В.С. Город как заказник: (пока?) неиспользуемая возможность // Памяти Германа Павловича Гапочки / Под ред. Д.Д. Соколова. М., 2018. C. 164–193.
Фридман В.С., Ерёмкин Г.С. Урбанизация «диких» видов птиц в контексте эволюции урболандшафта. М.: URSS, 2009. 240 s.
Фридман В.С., Ерёмкин Г.С., Захарова Н.Ю. Возвратная урбанизация – последний шанс на спасение уязвимых видов птиц Европы? // Russ. J. Ecosyst. Ecol. 2016. V.1. №4. DOI 10.21685/2500-0578-2016-4-3.
Хейр Б., Вудс В. Почему собаки гораздо умней, чем вы думаете. Спб: Питер, 2014. 145 с.
Шварц С.С. Экологические закономерности эволюции. М.: Наука, 1980. 153 с.
Шмальгаузен И.И. Избранные труды. Пути и закономерности эволюционного процесса. М.: Наука, 1983. 359 с.
Аgam Y., Vangel M., Roffman J.L. et al. Dissociable genetic contributions to error processing: a multimodal neuroimaging study// PloS one. 2014. V.9. №7. Р.e101784.
Alberti M., Correa C., Marzluff J. M. et al. Global urban signatures of phenotypic change in animal and plant populations// PNAS. 2017. V.114. №34. Р.8951-8956.
Alfimova M., Korovaitseva G., Lezheiko T. et al. Interaction effects of the COMT and DRD4 genes with anxiety-related traits on selective attention// Span. J. Psychol. 2014. V.17. P.e44. 9 pp.
Altink M.E., Rommelse N.N., Slaats-Willemse D.I. et al. The dopamine receptor D4 7-repeat allele influences neurocognitive functioning, but this effect is moderated by age and ADHD status: an exploratory study// The World J. Biol. Psych. 2012. V.13. №4. Р.293-305.
Armbruster D., Mueller A., Moser D.A. et al. Interaction effect of D4 dopamine receptor gene and serotonin transporter promoter polymorphism on the cortisol stress response// Behav. Neurosci. 2009. V.123. №6. Р.1288-1295.
Auerbach J.G., Faroy M., Ebstein R. et al. The association of the dopamine D4 receptor gene (DRD4) and the serotonin transporter promoter gene (5-HTTLPR) with temperament in 12-month-old infants// J. Child Psych. Psychiatr. 2001. V.42. №6. Р.777-783.
Babbitt C.C., Fedrigo O., Pfefferle A.D. et al. Both noncoding and protein-coding RNAs contribute to gene expression evolution in the primate brain// Genome biol. evol. 2010. V.2. P.67-79.
Bailey J.N., Breidenthal S.E., Jorgensen M.J. et al. The association of DRD4 and novelty seeking is found in a nonhuman primate model// Psych. Genet. 2007. V.17. №1. Р.23-27.
Barr C.S., Driscoll C. Neurogenetics of aggressive behavior: studies in primates// Neuroscience of aggression. 2013. V.17. P.45-71.
Baugh A.T., Witonsky K.R., Davidson S.C. et al. Novelty induces behavioural and glucocorticoid responses in a songbird artificially selected for divergent personalities// Anim.Beh. 2017. V.130. P.221-231.
Bhaduri N., Das M., Das A.B. et al. Dopamine receptor D4 exon 3 variable number of tandem repeat polymorphism: distribution in eastern Indian population// Ind. J. Human Gen. 2007. V.13. №2. Р.54.
Blumstein D.T. What chasing birds can teach us about predation risk effects: past insights and future directions// J. Ornithol. 2019. V.160. P.587–592.
Bonnefille R. Cenozoic vegetation, climate changes and hominid evolution in tropical Africa// Glob. Planet. Change. 2010. V.72. №4. Р.390-411.
Catalano M., Nobile M., Novelli E. et al. Distribution of a novel mutation in the first exon of the human dopamine D4 receptor gene in psychotic patients// Biol. Psych. 1993. V.34. №7. Р.459-464.
Chadaeva I.V., Bragin A.O., Kozhemyakina R.V. et al. Gene expression related to aggressive behavior on rat model// Bioinformatics of genome regulation and structure\systems biology (bgrs/sb-2018). The 11th Int.Conf. Abst. Novosibirsk: ICG SB RAS, 2018. P.203.
Chang F.M., Kidd J.R., Livak K.J. et al. The world-wide distribution of allele frequencies at the human dopamine D4 receptor locus// Hum. Gen. 1996. V.98. №1. Р.91-101.
Coyne S.P., Lindell S.G., Clemente J. et al. Dopamine D4 receptor genotype variation in free-ranging rhesus macaques and its association with juvenile behavior// Behav. brain res. 2015. V.292. P.50-55.
Cuvier G. Leçons d’anatomie comparée. Tome III. La première partie des organes de la digestion. Paris. Baudouin. 1805. 1-558 p., I-XXVIII p.
Cuvier G. Recherches sur les ossemens fossiles de quadrupèdes. Paris: Deterville. 1812. Т.1. 489 pp.
Delgado Jr R.A., van Schaik C.P. The behavioral ecology and conservation of the orangutan (Pongo pygmaeus): A tale of two islands// Evol. Anthrop. 2000. V.9. №5. Р. 201-218.
Diogo R., Molnar J.L., Wood B. Bonobo anatomy reveals stasis and mosaicism in chimpanzee evolution, and supports bonobos as the most appropriate extant mode l for the common ancestor of chimpanzees and humans// Sci.rep. 2017. V.7. №1. Р.1-8.
van Dongen W.F., Robinson R.W., Weston M.A. et al. Variation at the DRD4 locus is associated with wariness and local site selection in urban black swans// BMC Evol.Biol. 2015. V.15. №1. Р.1-11.
Dulawa S.C., Grandy D.K., Low M.J. et al. Dopamine D4 receptor-knock-out mice exhibit reduced exploration of novel stimuli// J. Neurosci. 1999. V.19. №21. Р.9550-9556.
Dulawa S.C., Grandy D.K., Low M.J. et al. Dopamine D4 receptor-knock-out mice exhibit reduced exploration of novel stimuli// J. Neurosci. 1999. V.19. №21. Р.9550-9556.
Ekelund J., Suhonen J., Järvelin M.R. et al. No association of the -521 C/T polymorphism in the promoter of DRD4 with novelty seeking // Mol. Psych. 2001 V.6. №6. Р. 618-619.
Fairbanks L.A., Way B.M., Breidenthal S.E. et al. Maternal and offspring dopamine D4 receptor genotypes interact to influence juvenile impulsivity in vervet monkeys// Psychol. Sci. 2012. V.23. №10. Р.1099-1104.
Falzone T.L., Gelman D.M., Young J.I. et al. Absence of dopamine D4 receptors results in enhanced reactivity to unconditioned, but not conditioned, fear// Eur. J. Neurosci. 2002. V.15. №1. Р.158-164.
Faurie C., Mettling C., Ali B.M. et al. Evidence of genotypic adaptation to the exposure to volcanic risk at the dopamine receptor DRD4 locus// Sci. rep. 2016. V.6. №1. Р.1-7.
Faurie C., Mettling C., Ali B.M. et al. Evidence of genotypic adaptation to the exposure to volcanic risk at the dopamine receptor DRD4 locus// Sci. rep. 2016. V.6. №1. Р.1-7.
Fruth B., Hohmann G. Food sharing across borders// Hum. Nat. 2018. V.29. №2. Р.91-103.
Gangi D.N., Messinger D.S., Martin E.R. et al. Dopaminergic variants in siblings at high risk for autism: Associations with initiating joint attention// Autism Res. 2016. V.9. №11. Р.1142-1150.
Garamszegi L.Z., Mueller J.C., Markó G. et al. The relationship between DRD4 polymorphisms and phenotypic correlations of behaviors in the collared flycatcher// Ecol. Evol. 2014. V.4. P.1466–1479.
Garroway C.J., Sheldon B.C. Urban behavioural adaptation// Mol.ecol. 2013. V.22. №13. Р.3430-3432.
Gelernter J., Kranzler H., Coccaro E. et al. D4 dopamine-receptor (DRD4) alleles and novelty seeking in substance-dependent, personality-disorder, and control subjects// The Amer. J. Hum. Gen. 1997. V.61. №5. Р.1144-1152.
Gazi L., Schoeffter P., Nunn C. et al. Cloning, expression, functional coupling and pharmacological characterization of the rat dopamine D4 receptor// Naunyn-Schmiedeberg’s arch. pharm. 2000. V.361. №5. Р.555-564.
Ghosh A., Seshadri M. Indian ethnic populations characterized by dopamine (D4) receptor VNTR polymorphism// Ann. Hum. Biol. 2005. V.32. №5. Р.574-584.
Gören E. The biogeographic origins of novelty-seeking traits// Evol. Hum. Behav. 2016. V.37. №6. Р.456-469.
Görlick M.A., Worthy D.A., Knopik V.S. et al. DRD4 long allele carriers show heightened attention to high-priority items relative to low-priority items// J. Cogn. Neurosc. 2015. V.27. №3. 509-521.
Goumas M., Boogert N.J., Kelley L.A. Urban herring gulls use human behavioural cues to locate food// Roy. Soc. open sci. 2020. V.7. №2. Р.191959.
Goumas M., Burns I., Kelley L. et al. Herring gulls respond to human gaze direction// Biology letters. 2019. V.15. №8. Р.20190405.
Grehan J.R., Schwartz J.H. Evolution of the second orangutan: phylogeny and biogeography of hominid origins// J. Biogeogr. 2009. V.36. №10. Р.1823-1844.
Gunbin K.V., Ponomarenko M.P., Suslov V.V. et al. Evolution of brain active gene promoters in human lineage towards the increased plasticity of gene regulation// Mol. Neurobiol. 2018. V.55. №3. Р.1871-1904.
Guan Z.H., Ma C.Y., Fei H.L. et al.. Ecology and social system of northern gibbons living in cold seasonal forests // Zool. Res. 2018. V.39. №4. Р.255-265
Hall B.K. Evolutionary Developmental Biology (Evo-Devo): Past, Present, and Future // Evolution: Education and Outreach. 2012. V.5. №2. Р 184–193.
Hare B., Wobber V., Wrangham R. The self-domestication hypothesis: evolution of bonobo psychology is due to selection against aggression// Anim.Behav. 2012. V.83. №3. Р.573-585.
Hejjas K., Vas J., Topal J. et al. Association of polymorphisms in the dopamine D4 receptor gene and the activity‐impulsivity endophenotype in dogs// Animal genetics. 2007. V.38. №6. Р.629-633.
Herron J., Freeman S. Evolutionary Analysis. NY: Pearson. 2014. 864 pp.
Hodgson N.W., Waly M.I., Trivedi M.S. et al. Methylation-related metabolic effects of D4 dopamine receptor expression and activation// Translat. Psychiatry. 2019. V.9. №1. Р.1-10.
Holmboe K., Nemoda Z., Fearon R.M.P. et al. Dopamine D4 receptor and serotonin transporter gene effects on the longitudinal development of infant temperament// Genes, Brain and Behav. 2011. V.10. №5. Р.513-522.
Holtmann B., Grosser S., Lagisz M. et al. Population differentiation and behavioural association of the two ‘personality’genes DRD4 and SERT in dunnocks (Prunella modularis)// Mol.ecol. 2016. V.25. №3. Р.706-722.
Inoue-Murayama M., Niimi Y., Takenaka O. et al. Allelic variation of the dopamine receptor D4 gene polymorphic region in gibbons// Primates. 2000. V.41. №4. Р.383-392.
Iofrida C., Palumbo S., Pellegrini S. Molecular genetics and antisocial behavior: where do we stand?// Exper. Biol. Medic. 2014. V.239. №11. Р.1514-1523.
Ito H., Nara H., Inoue-Murayama M. et al. Allele frequency distribution of the canine dopamine receptor D4 gene exon III and I in 23 breeds// J. Veter. Med. Sci. 2004. V.66. №7. Р.815-820.
Iwama H., Gojobori T. Identification of neurotransmitter receptor genes under significantly relaxed selective constraint by orthologous gene comparisons between humans and rodents// Mol. Biol. Evol. 2002. V.19. №11. Р.1891-1901.
James A.S., Groman S.M., Seu E. et al. Dimensions of impulsivity are associated with poor spatial working memory performance in monkeys// J. Neurosci. 2007. V.27. №52. Р.14358-14364.
Johnson M.T., Munshi-South J. Evolution of life in urban environments// Science. 2017. V.358. №6363. Р.eaam8327. https://science.sciencemag.org/content/358/6363/eaam8327
Keck T.M., Suchland K.L., Jimenez C.C. et al. Dopamine D4 receptor deficiency in mice alters behavioral responses to anxiogenic stimuli and the psychostimulant methylphenidate// Pharmac. Biochem. Behav. 2013. V.103. №4. Р.831-841.
Keltikangas-Järvinen L., Räikkönen K., Ekelund J., Peltonen L. Nature and nurture in novelty seeking// Mol. Psych. 2004. V.9. №3. Р.308-311.
Kitayama S., King A., Yoon C. et al. The dopamine D4 receptor gene (DRD4) moderates cultural difference in independent versus interdependent social orientation// Psychol. Sci. 2014. V.25. №6. Р.1169-1177.
Komiyama T., Iwama H., Osada N. et al. Dopamine receptor genes and evolutionary differentiation in the domestication of fighting cocks and long-crowing chickens// PLoS One. 2014. V.9. №7. Р.e101778.
Krützen M., Willems E.P., van Schaik C.P. Culture and geographic variation in orangutan behavior// Curr. Biol. 2011. V.21. №21. Р.1808-1812.
Kubinyi E., Vas J., Hejjas K. et al. Polymorphism in the tyrosine hydroxylase (TH) gene is associated with activity-impulsivity in German Shepherd Dogs// PLoS One. 2012. V.7. №1. Р.e30271.
Kühn K.U., Meyer K., Nöthen M.M. et al. Allelic variants of dopamine receptor D4 (DRD4) and serotonin receptor 5HT2c (HTR2c) and temperament factors: replication tests// Amer. J. Med. Gen. 1999. V.88. №2. Р.168-172.
Lakatos K., Toth I., Nemoda Z. et al. Dopamine D4 receptor (DRD4) gene polymorphism is associated with attachment disorganization in infants// Mol. Psych. 2000. V.5. №6. Р.633-637.
Lakatos K., Nemoda Z., Toth I. et al. Further evidence for the role of the dopamine D4 receptor (DRD4) gene in attachment disorganization: Interaction of the exon III 48-bp repeat and the− 521 C/T promoter polymorphisms// Mol. Psych. 2002. V.7. №1. Р.27-31.
Lakatos K., Toth I., Nemoda Z. et al. Dopamine D4 receptor (DRD4) gene polymorphism is associated with attachment disorganization in infants// Mol. Psych. 2003. V.5. №6. Р.633-637.
Lauzon N.M., Bishop S.F., Laviolette S.R. Dopamine D1 versus D4 receptors differentially modulate the encoding of salient versus nonsalient emotional information in the medial prefrontal cortex// J. Neurosci. 2009. V.29. №15. Р.4836-4845.
Laviolette S.R., Lipski W.J., Grace A.A. A subpopulation of neurons in the medial prefrontal cortex encodes emotional learning with burst and frequency codes through a dopamine D4 receptor-dependent basolateral amygdala input// J. Neurosci. 2005. V.25. №26. Р.6066-6075.
Lefebvre L., Aplin L.M. Social learning and innovation// Avian cognition. Chapt. 6. Cambridge: Cambridge Univ. Press, 2017. P.93-117.
Li J.J., Szkudlarek H., Renard J. et al. Fear memory recall potentiates opiate reward sensitivity through dissociable dopamine D1 versus D4 receptor-dependent memory mechanisms in the prefrontal cortex// J. Neurosci. 2018. V.38. №19. Р.4543-4555.
Liker A. Biologia Futura: adaptive changes in urban populations// Biologia Futura. 2020. V.71. P.1-8.
Livak K.J., Rogers J., Lichter J.B. Variability of dopamine D4 receptor (DRD4) gene sequence within and among nonhuman primate species// PNAS. 1995. V.92. №2. Р.427-431.
De Luca A., Rizzardi M., Buccino A. et al. Association of dopamine D4 receptor (DRD4) exon III repeat polymorphism with temperament in 3-year-old infants// Neurogenetics. 2003. V.4. №4. Р.207-212.
Ma X., Wang Zh., Zhang X. Evolution of Dopamine-Related Systems: Biosynthesis, Degradation and Receptors// J. Mol. Evol. 2010. V.71. №5-6. Р.374-384.
Maddox W.T., Gorlick M.A., Koslov S. et al. Serotonin transporter genetic variation is differentially associated with reflexive-and reflective-optimal learning// Cerebral Cortex. 2017. V.27. №2. bhv309.
Madrid J.E., Mandalaywala T.M., Coyne S.P. et al. Adaptive developmental plasticity in rhesus macaques: the serotonin transporter gene interacts with maternal care to affect juvenile social behaviour// Proc. Roy. Soc. B. 2018. V.285. №1881. Р.20180541.
Marchetto M.C., Hrvoj-Mihic B., Kerman B.E. et al. Species-specific maturation profiles of human, chimpanzee and bonobo neural cells// E-life. 2019. V.8. P.e37527.
Matsumoto M., Hidaka K., Tada S. et al. Polymorphic tandem repeats in dopamine D4 receptor are spread over primate species// Biochem. Biophys. Res. Comm. 1995. V.207. №1. Р.467-475.
McGeary J. The DRD4 exon 3 VNTR polymorphism and addiction-related phenotypes: a review// Pharm. Biochem. Behav. 2009. V.93. №3. Р.222-229.
McNew S.M., Beck D., Sadler-Riggleman I. et al. Epigenetic variation between urban and rural populations of Darwin’s finches// BMC evol. biol. 2017. V.17. №1. Р.1-14.
Miranda A.C. Mechanisms of Behavioural Change in Urban Animals: The Role of Microevolution and Phenotypic Plasticity// Ecol.Conserv. Birds in Urb.Envir. Eds. E.Murgui, M.Hedblom. Springer Int. Publ., 2017. Р.113–132.
Morrill K., Hekman J., Li X. et al. Ancestry-inclusive dog genomics challenges popular breed stereotypes// Science. 2022. V.376. №6592. Р.eabk0639.
Müller J.C., Carrete M., Boerno S. Genes acting in synapses and neuron projections are early targets of selection during urban colonization// Mol.Ecol. 2020. V.29. P.3403–3412.
Müller J.C., Edelaar P., Carrete M. et al. Behaviour-related DRD4 polymorphisms in invasive bird populations// Mol.Ecol. 2014. V.23. №11. Р.2876-2885.
Müller J.C., Edelaar P., Baños-Villalba A. et al. Selection on a behavior-related gene during the first stages of the biological invasion pathway// Mol.ecol. 2017. V.26. №21. Р.6110-6121.
Müller J.C., Partecke J., Hatchwell B.J. Candidate gene polymorphisms for behavioural adaptations during urbanization in blackbirds// Mol.Ecol. 2013a. V.22. №13. Р.3629-3637.
Müller J.C., Korsten P., Hermannstaedter C. et al. Haplotype structure, adaptive history and associations with exploratory behaviour of the DRD4 gene region in four great tit (Parus major) populations// Mol. Ecol. 2013b. V.22. №10. Р.2797-2809.
Nara H., Inoue‐Murayama M., Koshimura A. et al. Novel polymorphism of the canine dopamine receptor D4 gene intron II region// Anim. Sci. J. 2005. V.76. №1. Р.81-86.
Niimi Y., Inoue-Murayama M., Kato K. et al. Breed differences in allele frequency of the dopamine receptor D4 gene in dogs// J. Hered. 2001. V.92. №5. Р.433-435.
Nöthen M.M., Cichon S., Hemmer S. et al. Human dopamine D4 receptor gene: frequent occurrence of a null allele and observation of homozygosity// Human molec. genet. 1994. V.3. №12. P.2207-2212.
Owen R. Lectures on Comparative Anatomy and Physiology of the Invertebrate Animals, delivered at the Royal College of Surgeons in 1843. L.: Longman, Brown Green and Longman. 1843. 392 pр.
Paquet C., Portella A.K., Moore S. et al. Dopamine D4 receptor gene polymorphism (DRD4 VNTR) moderates real-world behavioural response to the food retail environment in children// BMC public health. 2021. V.21. №1. Р.1-9.
Pappa I., Mileva-Seitz V R., Szekely E. et al. DRD4 VNTRs, observed stranger fear in preschoolers and later ADHD symptoms// Psychiatry res. 2014. V.220. №3. Р.982-986.
Paterson A.D., Sunohara G.A., Kennedy J.L. Dopamine D4 receptor gene: novelty or nonsense? // Neuropsychopharm. 1999. V.21. №1. Р.3-16.
Patterson N., Richter D.J., Gnerre S. et al. Genetic evidence for complex speciation of humans and chimpanzees// Nature. 2006. V.441. №7097. Р.1103-1108.
Portella A.K., Papantoni A., Paquet C. et al. Predicted DRD4 prefrontal gene expression moderates snack intake and stress perception in response to the environment in adolescents// PloS one. 2020. V.15. №6. Р.e0234601.
Read D.W. Working Memory: A Cognitive Limit to Non-Human Primate Recursive Thinking Prior to Hominid Evolution // Evolutionary Psychology. 2008. Vol. 6, № 4. P. 676-714.
Reist C., Ozdemir V., Wang E. et al. Novelty seeking and the dopamine D4 receptor gene (DRD4) revisited in Asians: Haplotype characterization and relevance of the 2‐repeat allele// Amer. J. Med. Genet. B: Neuropsych. Genet. 2007. V.144. №4. Р.453-457.
Richards J.S., Arias Vásquez A., Von Rhein D. et al. Adolescent behavioral and neural reward sensitivity: a test of the differential susceptibility theory// Transl. Psych. 2016. V.6. №4. Р.e771-e771.
Riyahi S., Sánchez-Delgado M., Calafell F. et al. Combined epigenetic and intraspecific variation of the DRD4 and SERT genes influence novelty seeking behavior in great tit Parus major// Epigenetics. 2015. V.10. №6. Р.516-525.
Riyahi S., Björklund M., Mateos-Gonzalez F. et al. Personality and urbanization: behavioural traits and DRD4 SNP830 polymorphisms in great tits in Barcelona city// J. Ethol. 2017. V.35. №1. Р.101-108.
Ronai Z., Szekely A., Nemoda Z. et al. Association between Novelty Seeking and the −521C/T polymorphism in the promoter region of the DRD4 gene// Molecular psych. 2001. V.6. №1. Р.35-38.
Roussos P., Giakoumaki S.G., Bitsios P. Cognitive and emotional processing associated with the Season of Birth and dopamine D4 receptor gene// Neuropsych. 2010. V. 48. №13. Р.3926-3933.
Rubinstein M., Phillips T.J., Bunzow J.R. et al. Mice lacking dopamine D4 receptors are supersensitive to ethanol, cocaine, and methamphetamine// Cell. 1997. V.90. №6. Р.991-1001.
Schmidt L.A., Fox N.A., Hamer D.H. Evidence for a gene–gene interaction in predicting children’s behavior problems: Association of serotonin transporter short and dopamine receptor D4 long genotypes with internalizing and externalizing behaviors in typically developing 7-year-olds// Devel. Psychopath. 2007. V19. №4. Р.1105-1116.
Seaman M.I., Fisher J.B., Chang F.M. et al. Tandem duplication polymorphism upstream of the dopamine D4 receptor gene (DRD4)// Amer. J. Med. Genet. 1999. V.88. №6. Р.705-709.
Seaman M.I., Chang F.M., Deinard A.S. et al. Evolution of exon 1 of the dopamine D4 receptor (DRD4) gene in primates// J. Exper. Zool. 2000. V.288. №1. Р.32-38.
Selous F.C. Travel & Adventure in South-East Africa. Cambridge: Cambr. Univ. Press, 2011. 532 pp.
Seress G., Hammer T., Bókony V. et al. Impact of urbanization on abundance and phenology of caterpillars and consequences for breeding in an insectivorous bird// Ecol. Appl. 2018. V.28. №5. Р.1143-1156.
Serretti A., Macciardi F., Catalano M. et al. Genetic variants of dopamine receptor D4 and psychopathology// Schiz. Bull. 1999. V.25. №3. Р.609-618.
Shah A.A., Sjovold T., Treit D. Selective antagonism of medial prefrontal cortex D4 receptors decreases fear‐related behaviour in rats// Europ. J. Neurosci. 2004. V.19. №12. Р.3393-3397.
Shimada M.K., Inoue-Murayama M., Ueda Y. et al. Polymorphism in the second intron of dopamine receptor D4 gene in humans and apes// Biochem. Biophys. Res. Comm. 2004. V.316. №4. Р.1186-1190.
Sih A., Bell A., Johnson J.C. Behavioral syndromes: an ecological and evolutionary overview// Trend. ecol. evol. 2004. V.19. №7. Р.372-378.
Silva P.A., Trigo S., Marques C.I. et al. Experimental evidence for a role of dopamine in avian personality traits// J. Exp. Biol. 2020. V.223. №3. jeb216499
Spehar S.N., Sheil D., Harrison T. et al. Orangutans venture out of the rainforest and into the Anthropocene// Science Adv. 2018. V.4. №6. Р.e1701422.
Spelt A., Soutar O., Williamson C. et al. Urban gulls adapt foraging schedule to human‐activity patterns// Ibis. 2021. V.163. №1. Р.274-282.
Sprau P., Dingemanse N.J. An approach to distinguish between plasticity and non-random distributions of behavioral types along urban gradients in a wild passerine bird// Front. Ecol. Evol. 2017. V.5. №92.
Staes N., Guevara E.E., Helsen P. et al. The Pan social brain: An evolutionary history of neurochemical receptor genes and their potential impact on sociocognitive differences// J. Hum. Evol. 2021. V.152. P.102949.
Stavropoulos K.K., Carver L.J. Research review: social motivation and oxytocin in autism–implications for joint attention development and intervention// J. Child Psy. Psychiatr. 2013. V.54. №6. Р.603-618.
Strange B.A., Gartmann N., Brenninkmeyer J. et al. Dopamine receptor 4 promoter polymorphism modulates memory and neuronal responses to salience// NeuroImage, 2014. V.84. P.922-931.
Strobel A., Spinath F.M., Angleitner A. et al. Lack of association between polymorphisms of the dopamine D4 receptor gene and personality// Neuropsychobiol. 2003. V.47. №1. Р.52-56.
Suslov V.V. Par force evolution as a mechanism for rapid adaptation// Paleontological Journal. 2013. V.47. №9. Р.1048-1055.
Suslov V.V. Ontology of homologous series// BGRS/SB-2018. The 11th International Conference//Integrative Bioinformatics and Systems Biology (WIBSB-2018). First Sino-Russian Workshop. 22–23 August, 2018. Abstracts. Novosibirsk, ICG SB RAS, 2018. P. 61. DOI 10.18699/WIBSB-2018-46.
Tan T., Wang W., Williams J. et al. Stress exposure in dopamine D4 receptor knockout mice induces schizophrenia-like behaviors via disruption of GABAergic transmission// Schizophr. Bull. 2019. V.45. №5. Р.1012-1023.
Takeuchi H., Tomita H., Taki Y. et al. Cognitive and neural correlates of the 5-repeat allele of the dopamine D4 receptor gene in a population lacking the 7-repeat allele// Neuroimage. 2015. V.110. P.124-135.
Theofanopoulou C., Gastaldon S., O’Rourke T., et al. Self-domestication in Homo sapiens: Insights from comparative genomics// PloS Оne. 2017. V.12. №10. Р.e0185306.
Thompson M.J., Evans J.C., Parsons S. et al. Urbanization and individual differences in exploration and plasticity// Behav. Ecol. 2018. V.29. №6. Р.1415-1425.
Timm K., Tilgar V., Saag P. DRD4 gene polymorphism in great tits: gender-specific association with behavioural variation in the wild// Behav. Ecol. Sociobiol. 2015. V.69. №5. Р.729-735.
Timm K., Koosa K., Tilgar V. The serotonin transporter gene could play a role in anti-predator behaviour in a forest passerine// J. Ethol. 2019. V.37. №2. Р.221-227.
Trefilov A., Krawczak M., Berard J. et al. DNA sequence polymorphisms in genes involved in the regulation of dopamine and serotonin metabolism in rhesus macaques// Electrophoresis. 1999. V.20. №8. Р.1771-1777.
Tsuchimine S., Yasui-Furukori N., Kaneda A. et al. Minor genetic variants of the dopamine D4 receptor (DRD4) polymorphism are associated with novelty seeking in healthy Japanese subjects// Progr. in Neuro-Psychopharm. Biol. Psych. 2009. V.33. №7. Р.1232-1235.
Uzefovsky F., Shalev I., Israel S. et al. The dopamine D4 receptor gene shows a gender-sensitive association with cognitive empathy: evidence from two independent samples// Emotion. 2014. V.14. №4. Р.712.
Vandenbergh D.J., Zonderman A.B., Wang J. et al. No association between novelty seeking and dopamine D4 receptor (DRD4) exon III seven repeat alleles in Baltimore Longitudinal Study of Aging participants// Mol. Psych. 1997. V.2. №5. Р.417-419.
Vavilov N.I. The law of homologous series in variation // J. Genet. 1922. V.12. №1. P. 47—89. цит.по: Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. Л.: Наука, 1987. С.23-55.
Verhulst E.C., Mateman A.C., Zwier M.V. et al. Evidence from pyrosequencing indicates that natural variation in animal personality is associated with DRD4 DNA methylation// Mol.Ecol. 2016. V.25. P.1801–1811.
Wan M., Hejjas K., Ronai Z. et al. DRD4 and TH gene polymorphisms are associated with activity, impulsivity and inattention in Siberian Husky dogs// Anim. Genet. 2013. V.44. №6. Р.717-727.
Wang X., Pipes L., Trut, L. N. et al. Genomic responses to selection for tame/aggressive behaviors in the silver fox (Vulpes vulpes)// PNAS. 2018. V.115. №41. Р.10398-10403.
Watson H., Videvall E., Andersson M.N. et al. Transcriptome analysis of a wild bird reveals physiological responses to the urban environment// Sci. Rep. 2017. V.7. P.44180.
Wells T.T., Beevers C.G., Knopik V.S. et al. Dopamine D4 receptor gene variation is associated with context-dependent attention for emotion stimuli// Int. J. Neuropsychopharmac. 2013. V.16. №3. Р.525-534.
Wurth A. Behavior and genetic aspects of boldness and aggression in urban coyotes (Canis latrans). PhD diss., the Ohio State Univ., 2018. 192 pp.
Xie Z., Maddox W.T., Knopik V.S. et al. Dopamine receptor D4 (DRD4) gene modulates the influence of informational masking on speech recognition// Neuropsychol. 2015. V.67. P.121-131.
Zhang K., Grady C.J., Tsapakis E.M. et al. Regulation of working memory by dopamine D4 receptor in rats // Neuropsychopharmacology. 2004. V.29. №9. Р.1648-1655.
Zhong S., Israel S., Shalev I. et al. Dopamine D4 receptor gene associated with fairness preference in ultimatum game// PLoS One. 2010. V.5. №11. Р.e13765.
Zenner M.T., Nobile M., Henningsen R., et al. Expression and characterization of a dopamine D4R variant associated with delusional disorder// FEBS letters. 1998. V.422. №2. Р.146-150.
Zohsel K., Buchmann A.F., Blomeyer D. Et al. Mothers’ prenatal stress and their children’s antisocial outcomes–a moderating role for the Dopamine D4 Receptor (DRD4) gene// J. child psych. Psychiatry. 2014. V.55. №1. Р. 69-76.
Примечания
1Под ней понимается «перескок» редких видов из природных биотопов в урболандшафт (в особенности считающихся реликтами, стенотопными и т. д., не рудералов). Впервые я об этом узнал от А.Б.Чхобадзе в Вологде (см. пересказ его данных в «Глобальном экологическом кризисе…», с.175-178), а потом обсуждал её с Александром Нотовым из Твери на примере колонизации редкими видами откосов железных дорог (своего рода техногенный аналог сухих и мезофитных лугов, по их краю, в низине — высокотравных и заболоченных участков) или ботанических садов, см. статьи в приложении.
Подобные виды вдруг находят себе подходящее место в урболандшафте, расселяются, увеличивают численность, и именно на антропогенных участках, а не на остатках природных территорий внутри города, что совершенно контринтуитивно с точки зрения прежних представлений натуралистов о его биологии (опрокидываемых подобной урбанизацией). Данная ситуация сходна с интенсивно исследуемым ботаниками «убеганием» чужеродных растений из ботсадов и их распространением по городу, только это, наоборот, «прибегание» местных видов из окрестных лесов и лугов с тем же результатом.
2после появления человека разумного эти же изменения отмечаются в породной истории управляемых людьми собак и некоторых других одомашненных видов.
3А неотделимая от неё идея «самодоместикации бонобо» не работает вовсе.
4Это не значит, что различны и общие наборы генов, попавшие под отбор при доместикации и урбанизации. Наборы генов, демонстрирующих общий тренд отбора или дифференциальную экспрессию в этих двух случаях, могут перекрываться. Однако, во-первых, всегда есть специфичные гены, присутствующие в одном наборе и отсутствующие или слабо затронутые в другом. Во-вторых, общая практика сравнения полногеномных паттернов экспрессии показывает, что обычно различий здесь меньше, чем общего сходства. Что и привело к постгеномному кризису понятия гомологии.
5как минимум взрослых. Маленькие птенцы могут страдать от нехватки животных белков именно когда они необходимы вследствие достаточно частых случаев меньшего обилия голых гусениц и другого массового корма в городских биотопах [Seress et al., 2018].
6Аллелей с большим и малым числом повторов. Эта характеристика есть не у всех млекопитающих и часто отсутствует у птиц, но переносить результаты с одного вида на другой позволяет общая локализация повторов или полиморфизмов в экзон-интронной структуре гена DRD4, функции экзонов которого известны.
7Этим они сходны с людьми, у которых общая связь DRD4 с чертами индивидуальности может сильно (и что гораздо интересней, локально-обстановочно) варьировать у разных популяций и для разных черт. Связь перестраивается гибко и контекстуально, но нет её жесткого синдрома.
8В тесте на стратегию та не влияла на выигрыш, он был случаен.
9Это не неполное доминирование, иначе их стратегия была бы промежуточной.
10который давал бы преимущество равно в рисковой и нерисковой зонах.
11Волк, однако, из «конспецифика» стал главным врагом. У пастушьих собак отбор поощрял агрессию к «ближайшему на них похожему», «она подавлена лишь у пород, сформированных в отсутствие волков» (колли, шелти, бельгийская овчарка).
12Сходным образом у гиббонов подрода Hylobates DRD4 несёт от 4 до 8 повторов, но 4- и 5-повторные аллели отмечены лишь у островных видов [Inoue-Murayama et al., 2000].
13Чтобы его установить, нужны данные по гибридам, а их в статьях нет. С другой стороны, наследуемые признаки, ГР по которым строил Вавилов, имели друг с другом слабый эпистаз, даже если функционировали вместе (скажем, признаки цветов или плодов), ибо изменение радикала не влияло на признаки. Обратную ситуацию, ГР по признакам с сильным эпистазом исследовала Е.Н.Синская [1948].
14Именно так чаще всего «работают» клетки разных типов в тканях, или возникает зонарный окрас в отдельном волосе у собак либо в пере у птиц [Раутиан, 1987], что объясняет и Закон тканевого параллелизма А.А.Заварзина, и его связь с ЗГР Н.И.Вавилова. Объяснение ограничением пространства возможностей (ОПВ) для эволюции, предложенное самим А.А.Заварзиным, невалидно, хотя популярно в литературе [Раутиан, 1988]. Для органов, увязанных друг с другом по форме, топологии и функции, подобное ОПВ ещё уже, чем для тканей, однако к росту параллелизма это не ведёт. Первое знали уже Жорж Кювье [Cuvier, 1812] и Роберт Оуэн [Owen, 1843], второе — сам И.И.Шмальгаузен [1983], с его скепсисом к ЗГР. Последний, правда, не помешал современным эволюционистам объяснять ГР через теории Ивана Ивановича (не подходящие для этой задачи, см. главу 2.10). Г.С.Раутиан [1987] это делает даже после того, как для изменчивости аберраций окраски тетеревиных ей de facto пришлось вновь изобрести свободную комбинаторику субпризнаков окраса пера (ссылок на Вавилова здесь она не даёт) — иначе, по её же словам, ГР просто не строились. Проблема развития ЗГР — не подгон всех случаев параллелизма к одному объяснению с игнорированием исключений, а поиск условий, в которых работает каждое объяснение — нами подробно рассмотрена в главе 2.10 книги.
15Что равно типично как для примеров эпистаза в СТЭ, так и для контуров обратных связей, значимых в управлении физиологией индивида (объектов ЭТЭ). Так, мутация нефункциональности рецептора делает нефункциональным весь контур.
16объекты evo-devo варианта ЭТЭ [Herron, Freeman, 2014], СТЭ [Левонтин, 1978] и берговского номогенеза [Любищев, 1982].
17Разумеется, такое перекомбинирование отбором комплекса кофункционирующих и корегулируемых генов — не адаптивная сортировка особей в популяции, где элиминации может и не быть. При перекомбинировании работы комплекса генов элиминация неизбежна, а значит частоты аллелей изменятся. Но нет никакой гарантии, что это будет косвенный, регуляторный или стабилизирующий отбор — см. в главе 2.10. о функциональном перекрывании признаков.
18См. пересказ этих работ на русском: Александр Марков. «Ген авантюризма» чаще встречается у охотников-собирателей, чем у земледельцев.
19Этот порог удивительно однороден у разных человеческих обществ (обычно 40% суммы, у бывших советских людей 50%), что показывает нашу исходную приверженность равенству. См. «Животное и человеческое, расчёт и равенство»; «Людям свойственно стремление к равенству».
21Что впервые отмечено в трудовой теории происхождения человека Фридриха Энгельса и Людвига Нуаре, но социальный аспект здесь затмил биологический. При происхождении орангов Pongo spp. шёл подобный процесс, но остался незавершённым.
22Способностью жить и устойчиво действовать на фоне новизны — от дружелюбия через рисковость или невозмутимость к неофобии.
23Возможность такого отбора впервые показана М.М.Камшиловым: среда, ранее снижавшая у дрозофил выраженность мутации eyeless, стала её усиливать, связь признака со средой сохраняется вместо автономизации. Отбор здесь меняет лишь вектор, как для аллелей DRD4*5R у этносов Дальнего Востока сравнительно с европейскими.
24Впечатляющий “сдвиг” данного фенотипа с «длинного» аллеля на «короткий» как раз лучше объясним однотипным (снижающим) влиянием обоих на реакцию DRD4 с G-белком [Reist et al., 2007]. Вопреки этому сходству в биохимии, аллели противоположны по риску развития бредовой симптоматики у пациентов с тяжелыми психозами [Serretti et al., 1999].
25Скажем, нуклеотидная последовательность гена DRD4 или его метилирование, что прорисовывает связь последнего с урбанизацией у птиц [Riyahi et al., 2015] или сезоном рождения у человека [Roussos et al., 2010]. Однако у индивидуально-ориентированных субпризнаков поиска/исследования новизны таких связей не выявлено (см. дальше подробно обсуждаемую работу [Görlick et al., 2015]). Эффекты же полиморфизма разных районов гена DRD4 и его метилирования [Hodgson et al., 2019] комбинировать как раз могут, ибо опираются на разную биохимию передачи сигналов.
26Третий вариант — оптимизирующий (прежде всего зависимо от плотности) балансовый отбор между субпопуляциями одной популяции — подробно рассмотрен в литературе с точки зрения генетики [Левонтин, 1978] и её связей с эволюционной экологией или морфологией [Северцов, 2008], но вне связи с урбанизацией и не моделировался в доместикационном эксперименте. Поэтому для экономии места он не рассматривается, отметим лишь, что для урбанизации это препятствие [Фридман, Ерёмкин, 2009; Фридман и др., 2016], а генетико-популяционная теория, где какой-либо баланс необязателен, развита только Н.И.Вавиловым.
27Данный эффект блокировался анксиолитиками — фармакологическими средствами, снимающими тревогу, страх [Falzone et al., 2002]. У крыс, в отличие от мышей, блокада DRD4-антагонистами дала анксиолитический эффект [Shah et al., 2004] и ухудшила связь между безусловным и условным раздражителями в обонятельном условном рефлексе страха, [Laviolette et al., 2005]. Активация DRD4, напротив, усилила выраженность тех эмоциональных ассоциативных воспоминаний о страхе, которые в норме слабы, но не влияла на воспроизведение ранее заученных эмоционально значимых воспоминаний. Это блокировала активация гена DRD1 [Lauzon et al., 2009]. Видим, что блокада DRD4 и активация DRD1 равно создали крысам “слепую” (когнитивно-бесконтрольную) храбрость, однако она «cлепа» на разные субпризнаки памяти о страхе. Можно сказать, что блокада DRD4 сохраняет прошлый опыт, но мешает в нём разобраться, активация же DRD1 даёт автономию от прошлого опыта. Тогда нокаут по DRD4 просто мешает опыт приобретать, хотя и не блокирует такое приобретение [Tan et al., 2019, Falzone et al., 2002, Rubinstein et al.,1997], в том числе и у человека [Nöthen et al., 1994]. Опираясь на разные гены, оба субпризнака “слепой” храбрости могут по-вавиловски комбинировать в общем фенотипе (см. подробней главу 2.10 книги). С этим также хорошо согласуется влияние возраста на эффект DRD4 у такого долгоживущего и долгоучащегося вида, как наш собственный [Holmboe et al., 2011; Altink et al., 2012]. Учитывая рост СОПЖ в «городских» популяциях птиц и млекопитающих, это может проявиться и там (по крайней мере у крупных долгоживущих видов).
28Всем знакомый пример — поведение товарища Сухова в последнем бою “Белого солнца пустыни”. Убедившись в тщетности попыток сообщить появившемуся — неожиданно для всех, включая Сухова — Верещагину необходимость покинуть баркас или не заводить заминированный мотор, Сухов использует то обстоятельство, что в момент взрыва — неожиданного для всех кроме него! — друзья и враги равно взглянут на взрыв или хотя бы рефлекторно ослабят внимание (антисаккадная ошибка — неконтролируемый перенос взгляда на внезапно появившийся раздражитель с утратой контроля прочей обстановки [Agam et al., 2014]). На Сухова и даже в его сторону никто не будет смотреть, а обаятельного Верещагина всё равно уже не спасти. В момент взрыва Сухов, на ровном месте под носом у всех меняет огневую позицию, чем выигрывает бой. Итого, Суховым без потери фонового контроля над обстановкой (1) проведена её переприоритезация (2), в соответствие с чем снижена своя антисаккадная ошибка (3) — всё-таки взрыв, даже ожидаемый, очень сильный раздражитель, чтобы её полностью подавить — так, чтобы реализовать выбранный тип совладевания (бой с врагом) (4). Отвлечение внимания противника (5) и куда более интересный приём использования непроизвольного отвлечения самим противником своего же внимания своими же действиями (6) Сухов не раз демонстрирует в фильме. Последнее как раз спасает ему жизнь после незадачливого купания, окончившегося пленом.
29Действительно, это различие тем бледнее, чем больше в поведении рефлекторных и инстинктивных (по К.Лоренцу) составляющих и меньше рассудочной деятельности (по В.А.Вагнеру и Л.В.Крушинскому). Снижение вклада рефлекторной и инстинктивной компонент присуще именно особям в «городских» популяциях, увлечение ролью обратных связей и иных способов «связать» характеристики поведения в единый синдром — кибернетике, рефлексологии и сравнительной этологии. Неслучайно этот синдром разрушается при урбанизации вида.
30Пример из той же военной сферы: облака были повсеместной однозначной помехой для поиска самолётов противника прожектором, пока не додумались поднять истребители над облаками и подсветить последние по площадям: тогда и только тогда противник над облаками стал проецироваться на их фоне [Противовоздушная…, 1947]. Различение фона и помех, как и комбинаторика обмена между ними, происходящего здесь и сейчас, тесно смыкается с определением ряда в биологии, что впервые прослеживается в арабском (аль-Араби, ибн-Халдун), но не европейском естествознании. Последнее до сих пор не знает, что делать с ЗГР Вавилова и подозрительно косится на диалектику Г.Гегеля, толкуя её картезиански глобально, хотя она как раз создана для превращения помех в фон здесь и сейчас [Гегель, 1968-1973]. Нам так и не удалось найти в литературе дизайн опыта с комбинаторикой обмена фона и помех, происходящего здесь и сейчас, хотя, например, в литературе по эстетике возможный дизайн описан [Гегель, 1968-1973, Вёльфлин, 1999]. Поэтому о роли DRD4 в пункте III мы судим лишь качественно [Li et al., 2018]: для пп. I и II есть много вариантов количественного дизайна.
31В выборке [Görlick et al., 2015] превалируют представители этносов Западной Европы. В их подвыборке результаты перепроверены.
32Понятию «категории» данной работы в классификации К.Линнея соответствует существенный признак. Вообще проблема, на которую проливает свет это исследование, настолько важна для биологии и теории эволюции, насколько недооценивается биологами без полевого опыта. Скажем, сейчас популярна точка зрения, что из опытов по распознаванию целостного образа следует адаптивность генов-предикторов последнего, хотя животным в природе (и первобытному человеку) куда важней выявить замаскированного врага или добычу. Все охотники или путешественники-натуралисты отмечают, что детали (необычно движущиеся пятна — уши или конец хвоста) много важнее для выявления крупной кошки в засаде, чем её целостный образ [Корбетт, 2002; Selous, 2011; Арсеньев, 2022]. Второй автор наблюдал косулю у Кара-Дага, ориентируясь лишь на пятна «зеркала» и ушей. Дело было на посттуре конференции “Адаптационные стратегии живых систем”, и другие участники косулю не видели — все искали целостный образ, а не комбинировали пятна.
33При большем числе правильность решения задачи в: [Görlick et al., 2015] не отличалась от случайной.
348 одновременно говорящих были приравнены к энергетическим аудиопомехам [Xie et al., 2015].
35В поведении птиц в контексте урбанизации первое соответствует стратегии «медленного» исследования (глобальный поиск), второе — стратегии «быстрого» исследования (локально-обстановочная).
36Чего не позволял сделать дизайн опыта у верветок [Bailey et al., 2007]. В опыте с отложенным ответом с увеличением задержки высокоимпульсивные животные, независимо от генотипа по DRD4, давали меньше правильных ответов, чем низкоимпульсивные. Тем не менее, верветки-носители редких аллелей DRD4 имели тенденцию к более высокому обороту моноаминов в спинномозговой жидкости. Опять отмечаем гомологические ряды: импульсивность идентично влияет на поведение, но через биохимически различные механизмы [James et al., 2007].
37Об оппозиции индивидуализм vs коллективизм кульуры см. «Против «однониточных» объяснений». Сам признак «яркая выраженность» определим лишь как гомологическое сходство (по принципу, а не происхождению или функции). Действительно, изученные фенотипы дивергентны, повторы — сериальная гомология, а сходство биохимического модуля функции гена DRD4 [Reist et al., 2007] вообще трудно определить в рамках принятой иерархии сходств, где структурное (кювьерианское) сходство и филогенетическое родство понижаются в ряду от гомологии к конвергенции. Это ОПВ, но как его интерпретировать: оуэнова гомология? гомоплазия? параллелизм? С момента, когда Н.В.Тимофеев-Ресовский задался вопросом: гомологичны ли идентичные аминокислоты, закодированные разными диалектами или разными путями в связных сериях генетического кода? — проблема так и стоит перед любыми геномными сравнениями, оставаясь не разрешённой. Всё, что не гомология, по факту не различается, определяясь как сходство без родства, несмотря на обилие терминов. В такой ситуации невозможно сказать что-то определённое о причинах/условиях образования подобного сходства, хотя это важно эволюционной биологии [Суслов и др., 2019]. Решение было найдено в Законе гомологических рядов Н.И.Вавиловым, однако до сих пор остаётся в забвении, а сам закон во многом неверно интерпретируется.
38Несложно предположить, что успеху доместикации способствует превалирование коллективного исследования над индивидуальным, что и имеет место в действительности. Животное не выбирает хозяина. У.Рэли [1963] описывал индейских собак как бесхозных, готовых на контакт с любым числом людей, или одновременно (в загонной охоте) или по очереди (в роли мусорщика в посёлке). Индивидуально индеец охотился вообще без собаки.
39В такой ситуации оказываются и животные, совершающие нерегулярные перекочёвки по изменчивой мозаике ландшафтов: из наших ближайших родственников — антропоидов это не шимпанзе и гориллы, а орангутаны Pongo spp. и континентальные, но не островные виды гиббонов. На севере ареала биология этих последних изменяется к большему сходству с антропоидной: они живут увеличенными группами, формируют сеть убежищ-домициллу, включают в рацион регулярно листву, а эпизодически даже мясо древесных грызунов [Guan et al., 2018]. Африканские антропоиды в прошлом, как и теперь — стенобионты-эндемики: расселение орангов и гиббонов было прервано сначала трансгрессией океана, а дальше — хозяйственной деятельностью человека.
40Это в рафинировано-чистом виде помехоустойчивость-автономизация от среды, рассматриваемые как магистральный путь адаптивной эволюции в рамках эпигенетической теории эволюции (ЭТЭ): даже базовый интеллект и весь прошлый опыт оказались среди помех! Животные здесь автономизированы равно от внешнего окружения и внутренней среды воспоминаний. Правда, дизайн опыта не позволяет установить, идёт ли автономизация путём отрицательной обратной связи — ключевой признак классического варианта ЭТЭ (включающего теорию дестабилизирующего отбора Д.К.Беляева). Однако в других её вариантах [Филипченко, 1929; Hall, 2012; Махров, Болотов, 2019] достаточно просто помехоустойчивости.
41Вспомним, что город как биотоп для урбанизантов как раз не предполагает длительного пребывания в одной обстановке. Город — калейдоскоп непредсказуемой смены обстановок, и наоборот: возможность долгого обитания в одной обстановке, следования одной и той же поведенческой стратегии — ключевой признак для исходных, реликтовых, а часто и оптимальных [Махров, Болотов, 2019] видовых биотопов, почти никогда человеком не изменённых. Или таких биотопов, где выбора нет, как у домашних животных.
42По предположению Р.Roussos et al. [2010], за большую эмоциональную выносливость европейцы с «длинными» повторами DRD4 расплачиваются нарушением работоспособности, когда для принятия решения нужна эмоциональная обратная связь. Если так, то перед нами искомое разрушение регуляторного контура, отнюдь не ведущее к дестабилизации: все испытуемые были нормальными здоровыми добровольцами без проблем с психикой. Дело в том, что данное разрушение может быть комбинаторно скомпенсировано, а когнитивные механизмы принятия решений и планирования сохранены. Больше того, в повторяющейся ситуации высокой изменчивости среды нарушение этой обратной связи может купировать дистресс, защищать от беспокойства и депрессии, отвлекая внимание от эмоциональных невзгод [Roussos et al., 2010], а если рассудок расценивает хотя бы часть их как безопасные, то и обращая эти невзгоды в искомое удовольствие. См. выше работы о преобразованиях поведения и когнитивной сферы при урбанизации.
43Действительно, подростки с низким уровнем экспрессии DRD4 потребляли больше закусок, компенсируя дискомфорт от низкого социально-экономического статуса, чем повысили риск ожирения При высоком статусе тот же уровень экспрессии DRD4 ассоциирован с меньшим перееданием закусок [Portella et al., 2020]. Однако, зависимость небалансовая: в обоих случаях возможен и описан порочный круг.
44Чем сложнее изготовление орудий [Read et al., 2008] и выше сложность высказываний, тем важнее такая память, что прорисовывает связь полиморфизма DRD4, урбанизации, распознавания песен у птиц, речи и способности к языкам у людей. Вновь перед нами ГР похожести Вавилова — отбор на эти функции никак нельзя назвать конвергентным: конвергенция в эволюционной теории [Тимофеев-Ресовский и др., 1977] — cходство из-за отбора общим фактором среды (как вода для рыб и китов), аналогия — отбор общей функцией организма; здесь нет ни того, ни другого. См. подробнее главу 2.10.
45Сходный результат выявлен у верветок [Bailey et al., 2007, James et al., 2007], но здесь дизайн опыта не позволяет оба типа кратковременной памяти различить. Верветке надо просто вспомнить, в какой в ряду ящик положено лакомство, через время, заданное экспериментатором. С человеком (и только с ним) возможен более сложный дизайн: испытуемому кратковременно показывались строка букв и арифметический пример, затем — карточка с ответом на него (так что решать не обязательно, лишь прикинуть, верно или нет предложенное). А дальше испытуемым предлагали воспроизвести последовательность букв в строке: в каждом опыте приоритет у воспроизведения выше, но в сумме опытов должно быть не <85% верных решений примера [Görlick et al., 2015].
46Интересно рассмотреть гены, ассоциированные с такой памятью у людей, как кандидатные «гены урбанизации» для птиц и других позвоночных.
47В одном из них и лежит упомянутый SNP -521 C/T (rs1800955). Его связь с детекцией мозгом сигналов об ошибках и их коррекции для европейцев показана в случае обоих канонических маркеров ошибки — потенциала в ответ на её совершение и МРТ-активацию дорсальной передней части поясной извилины. Оба маркируют антисаккадную ошибку, но опираются на разную нейро- и генетическую основы [Agam et al., 2014].
Рекомендуем прочесть
 Пресечение урбанизации птиц как зеркало московского экоцида
Пресечение урбанизации птиц как зеркало московского экоцида Быстрая эволюция в связи с урбанизацией: примеры
Быстрая эволюция в связи с урбанизацией: примеры Сколько дятлов лесу нужно?
Сколько дятлов лесу нужно? Экоопасное расползание крупных городов срывает надежды на их устойчивое развитие
Экоопасное расползание крупных городов срывает надежды на их устойчивое развитие Урбанизация как основной фактор негативного влияния на местообитания диких животных Московского региона
Урбанизация как основной фактор негативного влияния на местообитания диких животных Московского региона Урбосреда как развивающая: сравнение с одомашниванием
Урбосреда как развивающая: сравнение с одомашниванием «Гены урбанизации» и приспособление к средовому стрессу
«Гены урбанизации» и приспособление к средовому стрессу Регенеративная урбанизация: Экополис
Регенеративная урбанизация: Экополис
