
Белобрюхая синица
Одна из основ сравнительной этологии — специфические результаты коммуникации созданы специфическими средствами, и первая «специфичность», в функции, детерминирована второй, в формах отдельностей, выступающих единицами поведения, ритуализованными демонстрациями. Выше это обосновано только косвенными данными, здесь приведены и прямые.
Дальневосточные белобрюхие синицы Parus minor, наряду с многими другими видами, имеют дифференцированную систему сигналов тревоги: выделенные этологами 3 крика дискретны, с альтернативными формами вокализаций (рис.1). Самый специфический (рис.1А) издаётся в связи с появлением змей — главной опасности для гнезда и птенцов.
Заслышав его при насиживании, синицы сразу же покидают гнездо, чтобы «змея» в дупле не застала врасплох, при воспроизведении вне дупла «автоматически» смотрят на землю рядом с гнездовым деревом. Т.е. сигнал вызывает защитное поведение, направленное именно против змей, отличающееся от защитного поведения при прочих опасностях, почему и относится к функционально-референтным (сравни с фактом 18). Всё это подтверждает специфичность действия сигнала, в противоположность издаваемому при появлении широкого круга хищников, птиц и млекопитающих (рис.1В).
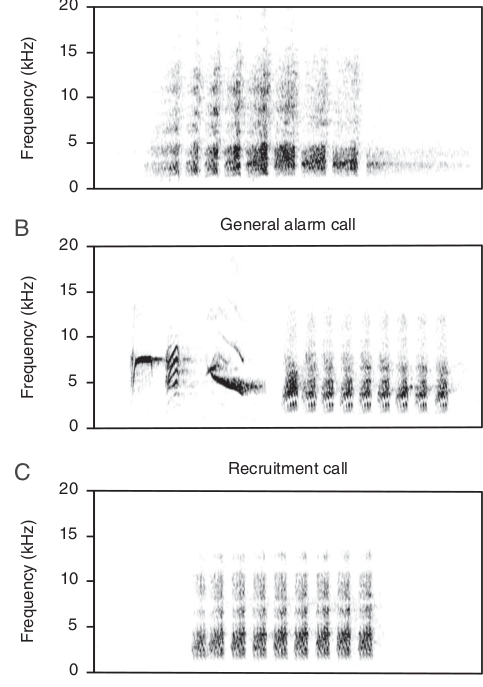 Рисунок 1. Спектрограммы криков тревоги белобрюхой синицы. А. Тревожный крик «на змею». В. Генерализованный крик тревоги. С. Призывный сигнал (привлечение сородичей в позитивном контексте, к гнезду или пище1). Ордината – частота в кГц.
Рисунок 1. Спектрограммы криков тревоги белобрюхой синицы. А. Тревожный крик «на змею». В. Генерализованный крик тревоги. С. Призывный сигнал (привлечение сородичей в позитивном контексте, к гнезду или пище1). Ордината – частота в кГц.
Toshitaka N. Suzuki (2018) исследовал, действительно ли тревожный сигнал «на змею» рождает специфический поисковый образ (англ. search image) именно данного хищника, облегчающий обнаружение именно данной опасности, а не не просто интенсифицирует осматривания? По примеру рождения мысленных (визуальных) образов в восприятии человеческой речи автор предположил порождение таких же при воздействии сигналов-символов и подтвердил это экспериментально.
Наличие поисковых образов подтверждает направлением внимания птицы в сторону наиболее вероятного появления именно данной опасности (появление прочих опасностей ожидалось бы в иных местах), и побуждением принимать за змею сколько-нибудь сходные с нею объекты, здесь — экспериментальную модель, деревянную палочку, перемещаемую так, что создавала впечатление ползанья. Иными словами, порождение поисковых образов значимо облегчает идентификацию опасности, направляя осматривания во вполне определённый район поля зрения и ускоряет определение объекта как «змеи», если он находится именно там и действительно похож на змею. Если это не так, проигрывание вокализаций рис.1А не улучшит обнаружение змееподобных объектов, лишь вызовет стереотипный ответ (скажем, взгляд вниз).
Всё это изучено T.Suzuki (2018) на свободноживущих белобрюхих синицах в начале сезона размножения. Схему опытов показывает рис.2. Проигрыванием криков тревоги «на змею» птиц привлекали к динамику, когда те оказывались в пространстве ~2 м над ним, им демонстрировали палочку длиной 18 см и диаметром 1,5 см. В эксперименте 1 её змееобразно двигали вверх по стволу соседнего дерева2 (рис.2А), в эксперименте 2 змееобразно двигали по земле (рис.2В) либо же двигали не змеевидно (раскачивали на низком кустарнике). Контроль — предъявление той же палочки в том же режиме при проигрывании генерализованных криков тревоги (рис.1В).
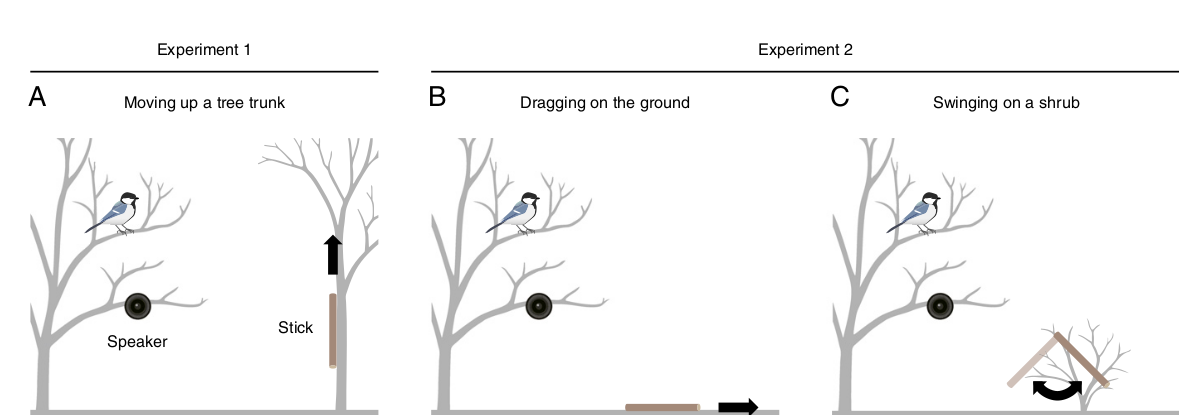 Рисунок 2. Схема экспериментов Suzuki, 2018. Объяснение в тексте.
Рисунок 2. Схема экспериментов Suzuki, 2018. Объяснение в тексте.
Если тревожный сигнал «на змею» рождает у птиц визуальный образ данной опасности, они именно и только в этом случае будут сильнее и специфичней реагировать на движение палочки — например, приближаясь к ней в случае змеевидных движений, но не иных. Это и происходило в действительности: крик «на змею» в 92% случаев (11 случаев из 12) побуждал белобрюхих синиц лететь к движущейся палочке: приблизившись на дистанцию <1 м, птицы осматривали окрестности (рис. 3А). Напротив, генерализованный тревожный сигнал заставил приблизиться на метр и менее только в 8% случаев ( (1 из 12; различия значимы по F-критерию: P < 0,001). В условиях выбора между двумя палочками, змеевидно движимой или нет (эксперимент 2), синицы, прослушавшие тревожный крик «на змею», устойчиво выбрали первую (10 из 12 тестирований, в 3-х приближение <0,5 м) и почти игнорировали вторую (1 приближение из 12, различия значимы по F-критерию, P < 0,0001). Генерализованный тревожный крик не привлёк внимания ни к одной из них (рис.3В).
Далее автор опроверг альтернативное объяснение, что разница реагирования вызвана тем, что сигнал «на змею» передаёт не визуальный образ опасности, а побуждение немедленно реагировать, генерализованный же тревожный сигнал нет.
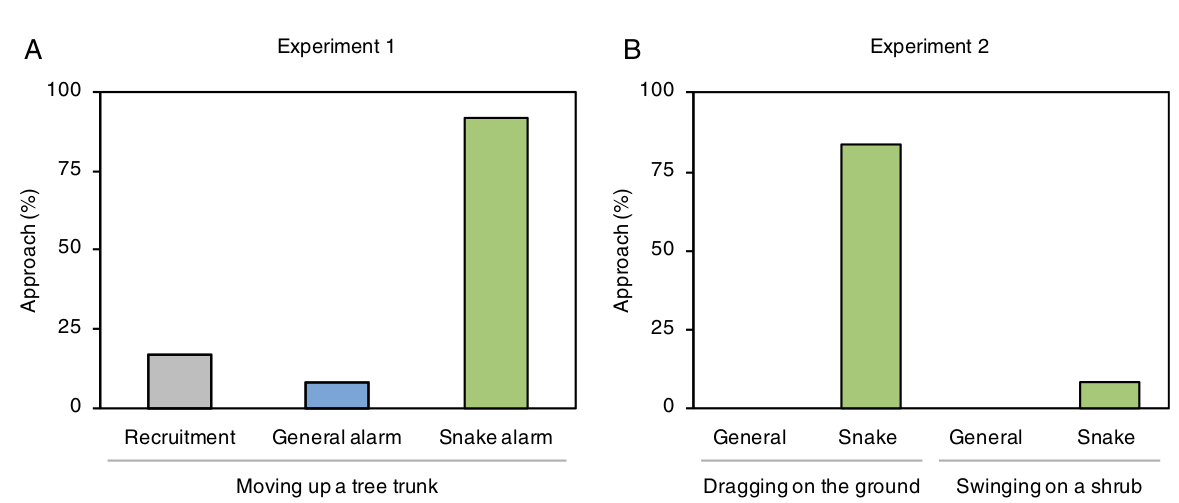 Рисунок 3. Результаты экспериментов 1 и 2, ордината — частота приближения к палочке, % (опыт 1 — движимой вверх по стволу, опыт 2 слева — движимой по земле, справа — раскачивающейся на кусту). Проигрываемые сигналы слева направо: опыт 1 — призывный крик, генерализованный крик тревоги, крик «на змею», опыт 2 — генерализованный и крик «на змею» в каждом из двух вариантов. Примечания. Каждую отдельную птицу каждым криком тестировали только однажды, в эксперименте 1 n=36, 2 — n = 48; поэтому n=12 для каждого из вариантов обоих опытов. Источник: Suzuki, 2018.
Рисунок 3. Результаты экспериментов 1 и 2, ордината — частота приближения к палочке, % (опыт 1 — движимой вверх по стволу, опыт 2 слева — движимой по земле, справа — раскачивающейся на кусту). Проигрываемые сигналы слева направо: опыт 1 — призывный крик, генерализованный крик тревоги, крик «на змею», опыт 2 — генерализованный и крик «на змею» в каждом из двух вариантов. Примечания. Каждую отдельную птицу каждым криком тестировали только однажды, в эксперименте 1 n=36, 2 — n = 48; поэтому n=12 для каждого из вариантов обоих опытов. Источник: Suzuki, 2018.
Однако при верности альтернативного объяснения приближение синиц к палочке при проигрывании обоих криков тревоги будет происходить значимо чаще, чем при обработке их нетревожными криками, например, призывным, который сам по себе не связан с каким-либо оборонительным поведением (рис.1С). Этого не наблюдается: синицы, слышавшие призывной крик, сближаются с палочкой даже несколько чаще слышавших генерализованную тревогу (рис.3А, 2 из 12 случаев, различия значимы по F-критерию). Эффекты обоих неспецифических криков значимо не отличаются от 0 (F-критерий, n=24, p=1,0).
Поэтому информированием о срочности реагирования различия в сближении с моделью змеи не объяснишь, лишь визуальным образом, созданным данным сигналом. Именно и только он толкает птиц искать поблизости «что-то похожее на змею», а увидев движение палочки по земле/по стволу, принять его за живую змею. Игнорирование палочки, двигающейся иначе, показывает специфичнось этого поискового образа — отнюдь не подробного, но «цепляющегося» за 1-2 ключевые черты (иначе бы птицы, не раз раньше встречавшие живую змею, игнорировали бы грубейшую модель их движения), как и «должно быть» у инстинктивного акта (факт 14).
Простые и красивые опыты Т.Сузуки фактически подтвердили схему реализации инстинкта, созданную отцам-основателями этологии (факт 5). Релизеры потому и зовут ключевыми раздражителями, что порождённые ими образы предельно схематичны, фактически сводятся к нескольким основным чертам, и вовсе не соответствуют образам, рождённым ментальными репрезентациями сознания человека или же антропоида, и даже этой же птицы, самостоятельно отслеживающей происходящее вокруг, без сигналов, канализирующих внимание и упрощающих картину.
После рождения этой схемы птицы активно отыскивают поблизости наилучшее соответствие ей и, найдя, реализуют инстинкт в виде сближения с палочкой, или атаки самцом колюшки модели, совсем непохожей, но красной снизу, или (в большинстве взаимодействий в природе) ответной демонстрации, позы или крика. В «грубости» поисковых образов, распознаваемых птицей, в автоматизме реагирования на сколько-нибудь соответствующие им объекты видим работу внутренних разрешающих механизмов, как их описывали при рождении сравнительной этологии, т.е. врождённого характера (факт 5).
Действительно, с настоящей змеёй синицы сближаются вплотную и зависают над ней, трепеща широко расправленными крыльями и хвостом: видимо, это «психологическая дуэль», когда обнаружение (а тем более «указывание» окрикиванием) стрессирует хищника, побуждая оставить атаку и уйти. Они никогда не вели себя так, приблизившись к палочке под действием сигнала: неспособные «уступить» вызванному им побуждению, поскольку оно инстинктивно, синицы непосредственно видели отсутствие искомой опасности, когда «бразды правления» поведением переходят к их собственной рассудочной деятельности, индивидуальному опыту и пр. Они и инстинкт находятся «в разных магистериях» (факт 3): услышав сигнал «на змею» внутри или вне гнезда, синицы реагируют по-разному, но это всегда оборонительное поведение, направленное именно против змей-разорителей гнёзд.

Японская московка
Действительно, тот же «поисковой образ» змеи появляется у японских московок P.ater insularis, прослушавших крик тревоги P.minor: вокализации близких видов гомологичны по форме (Suzuki, 2020). В опытах, также поставленных по схеме рис.2, московки после проигрывания чужих криков «на змею» значимо чаще сближались с палочкой, движимой змеевидно вверх по стволу (10 из 13 предъявлений) или по земле (7 из 13), чем движущейся иначе (раскачивающейся на кусту, 0 из 13), различия значимы по F-критерию, n=31, P<0,001 см. рис.4. Генерализованный крик тревоги или призывный крик белобрюхой синицы также были неэффективны (по 2 случая из 13 предъявлений для змеевидного движения по стволу/по земле, 0 случаев для раскачивающегося, результаты по F-критерию значимо не отличается от 0, n=24, p=1,0).
Поэтому приближение к палочке не объяснить подъёмом побуждения исследовать окрестности после прослушиваний резкого необычного крика другого вида (тогда б приближались к раскачивающейся палочке) ни разной скоростью обнаружения. Во втором случае московки, при проигрывании криков осматривающие верхнюю полусферу поля зрения, легче заметят палочку, движимую по дереву, смотрящие вниз — движимую по земле, и эта разница направлений сканирования может случайно совпасть с разницей в криках используемых для тестирования. Эта гипотеза опровергается отсутствием значимой связи между проигранным криком и пространственным расположением палочек или совместным влиянием этих двух переменных-регрессоров на вероятности приближения (отклик, обобщённая линейная модель регрессии, n = 56, χ2 = 0,22, df = 1, p = 0,64). Регрессионный анализ также показывает что на реакцию московок значимо влияют проигрываемые типы сигналов (n = 56, χ2 = 19,9, df = 1, p < 0,001), но не пространственное положение палочек (n = 56, χ2 = 0,82, df = 1, p = 0,37).
Действительно, межвидовое «подглядывание» и адекватное реагирование на сигналы другого вида распространено весьма широко (Magrath, Haff, Fallow и др., 2015), и интенсивно исследуется на тревожной и/или пищевой вокализациях (***). Ряд видов птиц воспринимают и точно используют для защиты от хищника информацию из тревожных криков млекопитающих (***), для которых это столь же обычно (***). Часть исследований доказывает адекватное «считывание» сигнальной информации из криков другого вида, отличающихся акустической формой, правда достаточно косвенные. Так, прекрасные расписные малюры Malurus cyaneus и белобровые кустовки Sericornis frontalis воспринимают информацию о степени неотложности реагирования на появившуюся опасность с воздуха, закодированную градуально (меньшая дистанция до опасных объектов побуждает добавить больше элементов в крик) в вокализациях обоих видов, и адекватно отреагируют её (Fallow & Magrath, 2010). Оба вида гнездятся в сходных местообитаниях, во внегнездовое время образуют смешанные стаи. Подчеркну: у обоих видов эти сложные крики «работают» как сигналы-символы, в противопоставлении сигналам «опасность с земли», с контрастирующей акустической формой.

Злато- и черношлемные калао
 Чëрно- и златошлемные калао, Ceratogymna atrata и Ceratogymna elata, демонстрируют «понимание» семантики дифференцированных сигналов-символов мартышек Диана, Cercopithecus diana. Они подлетают и окрикивают хищника, когда обезьяны издают крик «на орла», но не на «леопарда», ибо им угрожает только орёл (Rainey et al., 2004a, 2004b). Каролинские гаички замолкают и замирают (оборонительное поведение от ястреба) в ответ на крики «опасность с воздуха» американской хохлатой синицы, Baeolophus bicolor, но не другие тревожные вокализации вида. Однако они приближаются и участвуют в общем окрикивании хищника, заслышав вокализации уже делающей это группы синиц (Hetrick & Sieving, 2012).
Чëрно- и златошлемные калао, Ceratogymna atrata и Ceratogymna elata, демонстрируют «понимание» семантики дифференцированных сигналов-символов мартышек Диана, Cercopithecus diana. Они подлетают и окрикивают хищника, когда обезьяны издают крик «на орла», но не на «леопарда», ибо им угрожает только орёл (Rainey et al., 2004a, 2004b). Каролинские гаички замолкают и замирают (оборонительное поведение от ястреба) в ответ на крики «опасность с воздуха» американской хохлатой синицы, Baeolophus bicolor, но не другие тревожные вокализации вида. Однако они приближаются и участвуют в общем окрикивании хищника, заслышав вокализации уже делающей это группы синиц (Hetrick & Sieving, 2012).
Косвенность доказательств восприятия семантики тревожных сигналов другого вида следует из неточной (смещённой) оценки его адекватности именно и только по двигательному ответу: реакциям бегства (Magrath & Bennett, 2012), испуга (Carlile, Peters, & Evans, 2006) или моббинга (Templeton et al., 2005). В ряде случаев они больше указывают на уровень страха и стресса, чем на содержание воспринятой сигнальной информации (о типе опасности, иногда также о неотложности реакции на неё и пр.).
Более убедительны данные о направлении взгляда в ответ на разные сигналы репертуара, когда вид сталкивается с альтернативными типами опасностей (чаще всего «с воздуха» vs «c земли») и имеет в репертуаре «обозначающие» их дифференцированные сигналы, особенно сигналы-символы, вроде вышеописанных опытов T.Suzuki. Например, рыжеголовые Saguinus fuscicollis и усатые тамарины S.mystax дольше смотрели вверх в ответ на сигналы «опасность с воздуха», и вниз — в ответ на «опасность с земли3». Реакция на контрольный сигнал не показывает предпочтительного направления взглядов (их различали по поворотам головы). Реакция на проигрывание сигналов своего или другого вида сравнима с реакцией на реальную опасность и наступала много быстрее, чем на контроль (Kirchhof, Hammerschmidt, 2006).
У птиц поворот головы не так строго связан с направлением взгляда, как у приматов с их лицевым диском, но также полезен в анализе реакции на сигналы. Заслышав крики «опасность с воздуха», домашние куры поворачивали головы, вероятно, чтобы фиксировать её боковым зрением (Evans, Evans, & Marler, 1993). У трёх видов певчих птиц показан поворот клюва по направлению вероятной опасности после проигрывания тревожных сигналов, «указывающих» на альтернативные типы опасных объектов. При проигрывании крика «змея» сидящие на деревьях белобрюхие синицы направляли клювы к земле, заслышав же крики «опасность с воздуха», двигали головой в горизонтальной плоскости (Suzuki, 2012). Сидя на земле, австралийские сороки Cracticus tibicen после сигналов «опасность с воздуха» держали клюв вертикальней, чем после проигрывания генерализованного4 и смешанного тревожных сигналов (Kaplan & Rogers, 2013). Сидящие на ветвях медососы Manorina melanocephala в ответ на «опасность с воздуха» поворачивали клюв вверх, но держали горизонтально после проигрывания генерализованных криков (Farrow et al., 2017). Однако всё это качественные оценки, основанные на впечатлении наблюдателя, и неотделимые от риска ошибки.
1.
2.
3.
Рисунок 4. Сонаграммы альтернативных типов тревожных сигналов австралийской сороки (1) и шумного медососа (2), показаны первые 3,5 с. пятиминутной записи каждого из чётко дифференцированных криков. Обозначения. 1A — трель5, 1В — генерализованный крик тревоги, 1С — крик «опасность с воздуха». «Смешанный» крик тревоги в цитируемой работе составлен случайным чередованием В и С. Источник: Kaplan, Rogers, 20136. 2 — сонаграммы криков шумного медососа, воспринимаемых австралийскими сороками: A — «опасность с воздуха», издаётся на летящего хищника, В — «опасность с земли», побуждающий к моббингу, на наземного или сидящего пернатого хищника, С — крик красной розеллы Platycercus elegans (контроль). Обратите внимание на разность акустической структуры криков. 3 — измерение угла наклона клюва сороки, используемого как указатель направления поиска птицей потенциальной опасности. Источник: Pell et al., 2018.
Точное восприятие семантики коммуникативных сигналов другого вида было показано Pell et al., 2018, точно измеривших смену угла наклона клюва С.tibicen после проигрывания криков «опасность с воздуха» М.melanocephala, но не других криков, включая «опасность с земли» (рис.4). Кормясь там, сороки и манорины уязвимы для одного набора пернатых (ястреба австралийский бурый Аccipiter fasciatus и ошейниковый A.cirrocephalus, австралийский чеглок Falco longipennis, сапсан F.peregrinus, кукушечья иглоногая сова Ninox novaeseelandiae) и наземных хищников (лисы, собаки, кошки, змеи): все они активно окрикиваются обоими видами птиц.
Реакции сорок на вокализации манорин исследовали с января по июнь 2016 г. близ Канберры. Сначала определили, действительно ли угол наклона клюва (рис.4.3) отражает направление визуального поиска разнотипных объектов, в том числе потенциальных опасностей, фиксируя его изменения в ответ на движение мяча, брошенного на разной высоте или прокатанного по земле. При положительном ответе на этот вопрос угол наклона будет увеличиваться, когда птица видит бросаемый мяч в сравнении с прокатанным по земле, как он увеличивается7, когда они (или манорины) слышат крики «опасность с воздуха», тем более что в предыдущих работах (Kaplan, Rogers, 2013; Farrow et al., 2017) показана функциональная референтность данных криков обоих видов.
Оранжевый мяч диаметром 16 см катился по ровной земле 15-20 м или бросался в воздух так, чтобы он пролетел на расстоянии ~4 м от птицы. Мяч летел в воздухе примерно такое же расстояние и падал в 5-10 м за сорокой. Начало опыта маркировали тихим голосовым сигналом, записываемым на видео вместе со всем происходящим. Из 30 птиц 27 смогли протестировать обоими способами, видео, пригодные для измерения угла наклона клюва в обоих случаях, получены для 25. Действительно, наблюдая летящий мяч, сороки поднимают клюв выше, чем наблюдая катящийся по земле (рис.4), различия значимы. Смешанная линейная модель регрессии показывает, что угол наклона клюва детерминирован именно и только способом бросания мяча (χ2=39,74; Р<0,001), но не иными воздействиями, учитываемыми как случайные эффекты (присутствие молодых, пол, размер группы, порядок тестирования, индивидуальность птиц, взаимодействие способа бросания и порядка тестирования), см. Pell et al., 2018, table 1.
Рисунок 4. Угол наклона клюва сорок (левая ордината), наблюдающих мяч, летящий по воздуху (центральный столбец) и катящийся по земле (левый столбец). Правый столбец и правая ордината — разность углов наклона клюва в обоих случаях. Показаны средние значения, прогнозируемые на основании регрессии, и их 95% доверительный интервал.
Источник. Pell et al., 2018.
Далее птиц тестировали криками тревоги медососов, записанных в местах обитания тестируемых сорок, и вызванных предъявлением планирующей модели ястреба в натуральную величину. Её запускали с дистанции ~10 м от манорин и направляли с расчётом, что модель до приземления пролетит 5-10 м перед ними. Манорины издают крик «опасность с воздуха» на летящую модель, и переключаются на «опасность с земли» после её посадки. Их крики записывали в тех же местах, где позднее тестировали сорок, каждую отдельную птицу в не менее 100 м от предыдущей, чтобы понизить риск повторных записей одних и тех же птиц.
У австралийских сорок фиксировали угол наклона клюва при проигрывании трёх типов криков: «опасность с воздуха», «опасность с земли» манорин, либо контроль. Каждую из сорок протестировали криками «своего» медососа и розеллы, в первом случае всегда включавших 5 вокализаций, во втором от 4 до 8 (их подобрали и проигрывали так, чтобы громкость и длительность была одинаковой). Все 30 тестируемых сорок реагировали изменением угла наклона на тревожные крики манорин, и только 7 — на контроль (критерий Кохрана Q: Q2 = 46,0, P < 0,0001), поэтому далее им проигрывали лишь первые два. В ответ на «опасность с воздуха» они неизменно сильней поднимали клюв, т. е. искали опасность в более высоких секторах поля обзора, чем в ответ на «опасность с земли». В первом случае сравнительно со вторым значимо больше как максимальный, так и средний угол наклона клюва — на 7 и 6 градусов соответственно (рис.5). Регрессионный анализ показывает, что углы наклона клюва сорок, как средние, так и максимальные, т. е. направление поиска ими потенциальной опасности, определяются именно и только типом тестовых вокализаций (χ2 =10,091 и 14,272: Р<0,001), длительность всматривания — взаимодействием типа вокализаций и порядка следования обоих криков при проигрывании (χ2 = 5,025: Р<0,025). Влияния остальных факторов, образующих контекст реагирования на сигнал (размер группы, пол, доля молодых etc.), незначимы (Pell et al., 2018, table 2).
Длительность реагирования на крик «опасность с земли» не зависела от его предшествования крику «опасность с воздуха» или следования за ним, тогда как реакция на крик «опасность с воздуха» была дольше, когда он следовал за «опасность с земли», чем когда предшествовал ему (рис.6, Pell et al., 2018, table 2).
Рисунок 5. Максимальный (а) и средний (b) углы наклона клюва сорок (левая ордината) при воспроизведении криков «опасность с земли» vs «опасность с воздуха» манорин (левый и центральный столбцы). Показаны средние, прогнозируемые по регрессии table 2, и 95% доверительный интервал. Правый столбец и ордината — разность углов наклона клюва при реагировании на «опасность с воздуха» и на «опасность с земли» для каждой из тестируемых птиц. N = 30 сорок, протестированных обоими криками.
Рисунок 6. Длительность реагирования сорок в зависимости от порядка воспроизведения криков манорин: крик «опасность с воздуха» следует первым (левая пара столбцов) или вторым (правая). Столбцы — прогноз средних ± 95% доверительные интервалы; N = 30 сорок, по 15 на каждый порядок воспроизведения.
Количественный анализ реагирования, выполненный Pell et al., 2018, окончательно показал, что специфический результат коммуникации достигается специфическими средствами. Он был независимо (и не менее убедительно) подтверждён Jessica L. Yorzinski et al. (2013), использовавших ай-трекеры для прослеживания взглядов самок павлина при адресации ей роскошных комплексных демонстраций самца. На самок обрушивается множество разнородных воздействий, из которых показана значимость хохолка, числа глазков на шлейфе (это не хвост, а его верхние кроющие), длины самых длинных крайних перьев шлейфа, раздвоенных наподобие рыбьего хвоста; развёртывания шлейфа, устойчивость его удержания развёрнутым при движении, дрожи, пробегающая по настоящему (серому) хвосту.
Образы демонстраций токующего самца сменяют друг друга закономерно и предсказуемо (однотипно у одичавших павлинов Калифорнии и диких в Индии). Самка сближается с ним сбоку или сзади: завидев это, самец с определённой периодичностью трясёт приспущенными ярко-оранжевыми крыльями, также показывая хвост, и мелкими шажками пятится к самке (демонстрация сзади). Дальше он резко поворачивается на 1800 и наклоняет к ней перья шлейфа, словно охватывая её: перья переливаются, а дрожащий хвост шуршит и трещит, как листва под порывами ветра (фронтальная демонстрация). Самец старается сблизиться на дистанцию, меньшую длины шлейфа: тогда он интенсифицирует фронтальную демонстрацию, шелестит особенно сильно, и мелкими шажками движется к ней. При неготовности к спариванию он вновь поворачивается спиной, и демонстрации повторяются.
Главнейшая задача самца — привлечь и удерживать внимание самки, резко неодинаковый успех спариваний самцов связан именно с ним. Этологические данные показывают, что сила и специфичность внимания к видовым демонстрациям и другим стимулам определима по направлению взгляда, независимо от их «значения» и контекстов употребления, различающихся весьма и весьма. Авторы выучили павлиних носить беспроводные ай-трекеры, непрерывно фиксировавшие направление взгляда одного глаза, особенно области наиболее острого зрения, второй закрывали (рис.7). 6 таких самок успешно спаривались с демонстрировавшими перед ними самцами, 7 их отвергли, прочие не проделали ни того, ни другого, ибо самцы даже и не пытались: видимо, ношение окулографа не мешает обычному ходу взаимоотношений полов.
Рисунок 7. А. Самка с ай-трекером и закрытым вторым глазом. В. Кадр видео, показывающий работу датчика: зелёная точка — в данный момент самка смотрит на корпус, врезка справа вверху — датчик отслеживает положение её глаза и его поле зрения.
Рисунок 8. Шанс обращения внимания самки на разные составляющие образа самца (1-10) в моменты фронтальной демонстрации (A) и демонстрации сзади (B, C). Обозначения. Белое — шансы выше случайного уровня, жёлтое — ниже, зелёное — ему соответствует; 1 — верхние раздвоенные перья шлейфа, 2 — верхние глазки, 3 — нижние раздвоенные перья шлейфа, 4 — нижние глазки, 5 — плотные перья внизу шлейфа, далее в А 6 — хохолок с головой, 7 — чешуеобразные перья, 8 — корпус и 9 — ноги самца; в B-C 6 — хвост; 7 — белые и 8 — черные перья; 9 — крылья; 10 — ноги.
Авторы изучили дистанции, с которых самцовые демонстрации привлекают внимание самок, и степень специфичности взгляда – он схватывает целенаправленно отдельные компоненты образа (скажем, глазки, раздвоенные перья шлейфа), или же холистичен и пробегает фигуру в целом? Прежде всего, павлинихи точно направляют взгляд на несоциальные стимулы, вроде корма, бросаемых перед ними мелких предметов или потенциальной опасности (чучело енота) с минимальной ошибкой (3,90±0,39°, диапазон 1,75–5,87°, N=14). В 15.9% случаев ошибок они совершали корректирующие саккады, чтобы направить взгляд точно на стимул: цепко держали внимание именно на предметах, и значимо чаще взгляд останавливали на них, чем на той же области до демонстрации.
Самки павлинов следят за адресованными им демонстрациями не более трети времени (27,5% при фронтальной, 21,3% при демонстрации сзади), чем создают впечатление собственной незаинтересованности. Это дезориентировало Е. Н. Панова, увидевшего ещё одно подтверждение своей убеждённости в неспецифичности демонстраций и их семантической пустоте8. Однако на павлиних действует вовсе не образ в целом (как было бы при его неспецифическом действии, лишь возбуждающем самку); их взгляд фиксирует строго определённые части демонстраций, причём различающиеся в случаях, когда они видят её вблизи или издали.
Оказывается, самочьи взгляды «схватывали» строго определённые компоненты образов демонстраций, отчасти сходные у обоих, отчасти различающиеся, и игнорировали другие (рис.8, белый и жёлтый). Первые отличают это (коммуникативное) внимание от всякого прочего, направленного, скажем, на потенциальные опасности или других самцов, видимых одновременно с токующим; вторые показывают различение демонстраций их адресатками. При предъявлении фронтальной демонстрации самки чаще случайного фиксировались внизу образа — на нижних глазках, раздвоенных перьях шлейфа, густых перьях ниже него и ногах, за исключением крыльев; и реже случайного они смотрели на верхнюю половину образа, хохолок и чешуйчатые перья (рис.8). Наблюдая демонстрацию сзади, самки чаще случайного смотрели на крылья самца и те же самые частями тела, что при демонстрации спереди — глазки нижней части шлейфа, раздвоенные и густые перья. Реже случайного их взгляд останавливался на ногах, чёрных и белых перьях, и тех же компонентах верхней части образа, что во фронтальной демонстрации. В обоих случаях взгляды самок «ходили» справа налево и наоборот, охватывая по всей ширине образ, репрезентируемый самцом (рис.9).
Рисунок 9. Движение взгляда трёх разных самок, оценивающих демонстрации спереди (А-С) и сзади (D-F). Размер чёрных кружков пропорционален времени рассматривания соответствующих частей образа, представляемого самцом.
Возможно, так самки оценивали развитость и симметричность брачных украшений самцов: раньше было показано, что они производят «отсечение снизу», избегая партнёров с минимумом глазков. Также возможно, что ширина шлейфа, оцениваемая такими взглядами, коррелирует с его длиной, значимой для выбора самок в ряде павлиньих популяций.
Верхнюю часть шлейфа с его глазками самки обычно рассматривают около 5% времени. Однако в эксперименте с искусственным шлейфом, нижнюю часть которого скрыли, павлинихи не просто дольше смотрели на верхнюю, но значимо дольше, чем контрольная группа самок — на верхнюю часть ветвей сравнимой длины (Jorzinsky et al., 2013, table 1). Ещё важнее, что самки, увидевшие подобный шлейф издали, начинали сближение с ним с меньшей задержкой, чаще рассматривали его и, приблизившись, дольше пробыли рядом, чем при показе ветвей сравнимой длины. Поэтому если низ шлейфа — очевидный контактный сигнал, верх может функционировать быть дистантным сигналом, также как нижняя очевидный контактный.
Далее авторы выявили управляемость взглядов самок демонстрированием самца, и наоборот — смена им демонстраций зависит от внимания самок. Ранее аналогичное «управление» развёртыванием брачных демонстраций самца со стороны самки они установили у атласного шалашника, используя самок-роботов9. Павлинихи значимо дольше смотрели на фронтальную демонстрацию, когда в её рамках хвост самца шелестит (лишь в 28,3% случаев), чем без этих звуков (51,2±2,4% vs 37,4±2,2). Они также дольше смотрели на демонстрацию сзади, когда в её рамках самец тряс крыльями (64,4% случаев), чем при беззвучном исполнении (33,16±2,8% vs 15,0±2,3% Jorzinsky et al., 2013, table 1).
Данные формы поведения — единственные звучащие компоненты демонстраций. Анализ направления взглядов самок до, после и во время названных демонстраций показывает, что их привлекли именно шелест и звук встряхиваемых крыльев, а не что-то еще (рис.10). Демонстрация сзади (но не спереди) обладает и последействием: при её исполнении самки больше рассматривали репрезентируемые им образы самцов не только в момент шелестения или тряски крыльев, но и потом (рис…). В отличие от них, само по себе уменьшение дистанции не вызывало таких изменений взгляда.
Рисунок 10. Среднее (± стандартная ошибка средней) время рассматривания самками фронтальной демонстрации (А) и демонстрации сзади (В) до, во время и после исполнения их звучащих компонентов, % общего времени рассматривания. N=11 самок (А) или 9 самок (В).
Минимизируя дистанцию до самки, самцы привлекают к себе больше внимания. Самки дольше смотрели на демонстрацию сзади при их движении, чем при нахождении на месте, 31,3±3,2% vs 25,6±2,1%, и на демонстрацию спереди, когда самцы были в 1-2 м от них, чем в 3-4 и более, 45,5±2,2% vs 35,3±3,6%), однако большая тщательность взгляда увеличивает риск быть отвергнутым самкой.
Выяснилось, что реакция самки не только специфична, но и последовательна. Сначала внимание самки «ловится» и удерживается наиболее быстрыми и изменчивыми движениями (на которые и мы первыми обращаем внимание) — шелест шлейфа и трепетание крыльев. А вот оценки самца и «оргвыводы из неё» — сближаться с ним вплоть до спаривания или нет — зависят от степени стереотипности экспозиции шлейфа со всеми сопровождающими эффектами, визуальными и акустическими. Верхняя часть шлейфа привлекает внимание на дальней дистанции и побуждает приблизиться, нижняя оценивается самкой вблизи. Соответственно, множественное дублирование сигналов в коммуникации животных (котором много и правильно пишет Е.Н.Панов) осуществляется способом, прямо противоположным его представлениям — отнюдь не за счёт того, что разные компоненты мультимодального сигнала равно пусты семантически, но прямо наоборот, за счёт того, что разные компоненты последовательно и специфически управляют поведением реципиента. Когда (или если) сработали одни, в ход вступают другие и пр. Всё это даёт новую жизнь иерархической концепции инстинктов Нико Тинбергена, поскольку разные демонстрации — сериальные компоненты инстинкта, как членики у кольчатого червя, организованный обмен ими во взаимодействиях, т.е. коммуникация – кооперативная реализация особями какого-либо инстинкта, изоморфная совместному исполнению танца.
Эти данные ликвидируют большую часть критики Е.Н.Панова в адрес полового отбора, собранной в недавно вышедшей книге «Половой отбор: теория или миф?». Таки да, как и у ткачика-вдовушки, самки привлекаются лучшими брачными украшениями, с учётом того, что существенна не развитость их сама по себе, а стереотипность исполнения демонстраций, предъявляющих «действующий образ» самке. Это и есть половой отбор, вполне реально существующее явление, также как отбор в конкуренции за территорию, социальный статус и другие формы социального отбора. Даже господь бог не сделает сущее небывшим; а вот что можно и нужно критиковать, это попытки увидеть половой отбор везде и всюду на волне соответствующей моды, и социобиологические представление об асимметричности этого процесса, когда из-за якобы меньших затрат самцы максимизируют спаривания, а самки — привязанность самцов.
Примечания
1Может использоваться для мобилизации на окрикивание хищника, если Р.minor издают этот крик сразу вслед за специфическим криком тревоги: «точность обозначения» опасности усиливает готовность тратить время и силы на моббинг.
2В обоих случаях её протягивали вперёд на тонкой нити. В районе исследований никто, кроме змеи-разорителя гнёзд островного полоза Elaphe climacophora не движется по стволам таким образом.
3В ответ на опасность с воздуха обезьянки смотрят вверх, и дальше спускаются пониже; заметив её на земле, смотрят вниз и приближаются к ней.
4Издаётся в ответ на наземных хищников, побуждает приблизиться и окрикивать. В смешанной ситуации может чередоваться с криком «опасность с воздуха», поэтому в описываемых исследованиях фигурирует «смешанный крик тревоги», составленный из обоих типов вокализаций. При первом появлении воздушного хищники сороки всегда издают лишь крик «опасность с воздуха»: смешанный крик тревоги и другие вокализации отмечаются позже, когда (или если) поведение птиц меняется, есть возможность приблизиться к нему и окрикивать, нет императивности немедленного спасения жизни.
5Используется как подпесня, звучит при обучении молодых песне взрослых самцов, при отдельном употреблении — сигнал социальной аффилиации или пищевой. Все контексты употребления трели неизменно неагрессивные и не связанные с охраной территории.
6Особенная прелесть этой работы — наряду с сугубой специфичностью ответа сорок на вокализации «опасность с воздуха» (проигрывание этих, но не других криков устойчиво обращает взгляд вверх во время и отчасти после, но не до тестирования) показана стабильность реагирования на этот крик в географически разных популяциях (города и сельская местность Нового Южного Уэлльса, о.Тавеуни из архипелага Фиджи, куда сорок завезли и где они изолированы >10 поколений). Смешанный крик действует слабее крика «опасность с воздуха», как и должно быть при комбинировании релизера с поведением, незначащим в данной ситуации. В сельской местности, где риск гибели от пернатых хищников выше, реакция сорок на этот крик сильнее и специфичней, чем в городе. Во всех трёх местностях сорокам угрожают разные пернатые хищники, риск гибели от них варьирует у птиц разного возраста и состояния, но все они одинаково специфически «обозначены» этим криком (Kaplan, Rogers, 2013). Его семантика относится к типу опасности, но не к варьирующим обстоятельствам возможного нападения: последние (например, неотложность реакции, заданная дальностью до вдруг появившихся опасных объектов) могут «обозначаться» другими криками или деталями издавания данного. Во всех этих случаях неизменно устойчива и взаимно-однозначна связь означаемого и означаемого сигнала, пусть не всегда легко обнаруживаемая.
7По мнению наблюдателя, чьё впечатление бывает ошибочно. Достоинство данной работы в точном количественном определении изменений угла наклона, где данные видеосъёмки и двойного слепого контроля оценивания угла наблюдателем проверяют и уточняют результаты друг друга. Тестировали 30 сорок (по 15 самцов и самок): у каждой мерили изменение угла наклона клюва сперва в ответ на летящий по воздуху мяч в сравнении с катящимся по земле, затем в ответ крик манорин «опасность с воздуха» в сравнении с «опасность с земли». Опыты ставили в сухие дни, с небольшим ветром, в отсутствие помех в виде территориальных конфликтов или хищников и манорин поблизости. Сорок привлекали сыром и после >1 мин спокойного питания тестировали, фиксируя размер группы и наличие молодых особей как возможные помехи в реагировании. Прочие птицы считались «присутствующими» при нахождении ближе 5 м к тестируемой.
8 См. Панов Е.Н. Сведения о брачном поведении индийского павлина. Вводные замечания к переводу статьи Takahashi M., Arita H., Hiraiwa-Hasegawa M., Hasegawa T. (2008) Самки индийского павлина не отдают предпочтений самцам с наиболее эффектным шлейфом, https://journal.panov-ethology.ru/img/1-2012/Pavlin.pdf
Подробно описывая брачные демонстрации павлина (в том числе собственные впечатления от наблюдений за ними в «Аскании-Нова») в обоснование отсутствия полового отбора и/или специфичности действия демонстраций, Евгений Николаевич неизменно отказывается цитировать эту и другие работы, противоречащие его представлениям о коммуникативном поведении и демонстрациях позвоночных (во многом сформировавшихся априори). Что сильно обесценивает его аргументацию: цитировать одни подтверждения собственным взглядам – понятная человеческая слабость, особенно у исследователей, заслуженно признанных лидером данного направления, однако неизменно ведущая к досадным ошибкам, и эта история не исключение.
9 См. «Специфичность мотивационной подосновы демонстраций и их семантичность показаны строго», https://www.socialcompas.com/2025/02/21/spetsifichnost-motivatsionnoj-podosnovy-demonstratsij-dokazana/ Выяснилось, что структура последовательности брачных демонстраций самца регулируется ответом самки — именно стремлением избежать разрыва в её коммуникативном внимании, чреватого тем, что она «разочаруется и уйдёт» (вызвав нешуточный стресс обоих, особенно у самца). Самец, демонстрируя, не «разогревает» самку, а «устанавливает контакты» и далее «развивает диалог», последовательно меняя демонстрации, «наращивая планку» ритуализированного ухаживания. Отсюда обязателен вывод о «семантичности» демонстраций, «работающих» предикторами поведения обоих участников взаимодействия.
Рекомендуем прочесть
 Специфичность мотивационной подосновы демонстраций и их «семантичность» доказаны строго
Специфичность мотивационной подосновы демонстраций и их «семантичность» доказаны строго Опять о феромонах
Опять о феромонах Используются ли «понятия» в коммуникации животных?
Используются ли «понятия» в коммуникации животных? Про «фантомный инстинкт»
Про «фантомный инстинкт» Коэволюция парохиального альтруизма и войны
Коэволюция парохиального альтруизма и войны Поведенческие изменения в «городских» популяциях птиц — нет общего с одомашниванием
Поведенческие изменения в «городских» популяциях птиц — нет общего с одомашниванием Вольфганг Шлейдт. Коммуникация животных. Поиски понимания сложности поведения
Вольфганг Шлейдт. Коммуникация животных. Поиски понимания сложности поведения Миролюбивая культура среди диких павианов: её возникновение и передача
Миролюбивая культура среди диких павианов: её возникновение и передача
